





This chapter presents examples of the pragmatic application of the newly uncovered capabilities. On this basis, it confirms the necessity for profound changes in diagnostic and therapeutic approaches in dermatology—both as a scientific discipline and as a clinical practice.
— A. Teslinov
Scientific activity, through the methodology of cognition, considers the real picture of the world and serves as a primary means of transforming scientific knowledge into practice. This transformation is realized through the invention of methods of scientific investigation and their application.
The selection of markers that differentiate cellular subpopulations and characterize the dynamics of membrane events within these subpopulations made it possible to obtain a series of quantitative and functional assessments of skin cells in the volunteers who participated in the study.
After sample preparation as described above, the number of viable skin cells in volunteer A., 42 years old, was assessed by flow cytometry (Fig. 36). The result was 94%.
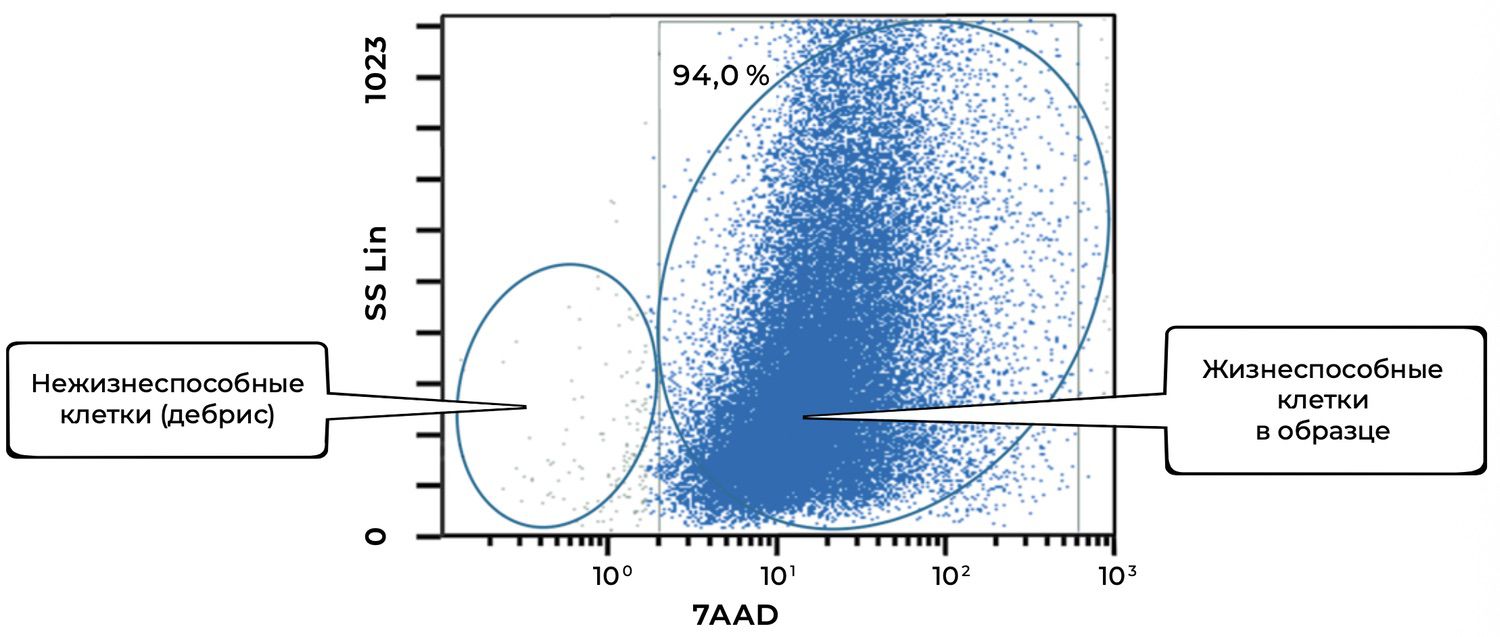
Figure 36. Relative number of all viable (7AAD⁻) cells in the skin-sample suspension, determined by flow cytometry
The overview cytoimmunogram (Fig. 36) shows the assessment of viability of skin cells obtained from the biopsy sample. The purpose of the analysis was to determine the proportion of viable cells within the total cellular population using the 7AAD dye, which penetrates only cells with damaged membranes (i.e., non-viable cells), and the SS Lin (side scatter) parameter, which reflects preservation of intracellular structure.
X-axis — 7AAD (log scale).
Higher values correspond to membranes permeable to the dye → non-viable cells.
Y-axis — SS Lin.
Reflects granularity and internal structural complexity of the cell.
Identified populations
Viable cells (right cluster, 94.0%)
Description: Cells with intact membranes, not stained by 7AAD (low fluorescence on the X-axis).
Parameters: A bright, dense cluster with broad heterogeneity in SS Lin, reflecting the diverse cell types of the skin.
Conclusion: The vast majority of cells in the sample are viable.
Non-viable cells / cellular debris (left cluster)
Description: Cells with disrupted membranes or fragments, strongly stained by 7AAD.
Parameters: Located in the lower-left region of the plot, with low granularity and high membrane permeability.
Thus, the total proportion of viable cells in the sample is 94.0%.
This indicates excellent sample quality and suitability for downstream phenotyping, population analysis (keratinocytes, fibroblasts, etc.), or cell-based applications.
The low proportion of debris reflects appropriate sample handling and gentle cell isolation.
In addition, specific cell phenotypes were quantified using panels of fluorochrome-labeled monoclonal antibodies targeting defined membrane receptors. The sample contained 54.2% keratinocytes, of which 41.4% were in an activated state (Fig. 37).
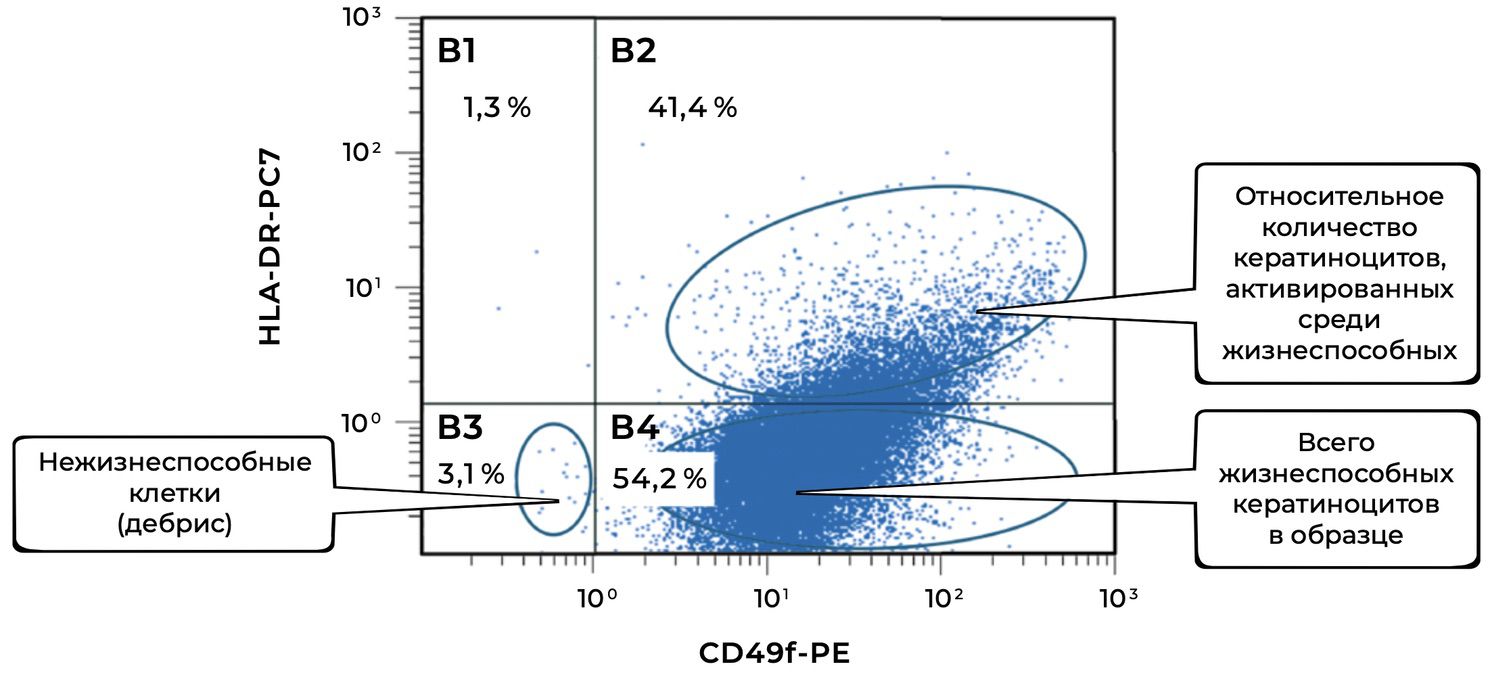
Figure 37. Ratio of viable keratinocytes (CD49f⁺ phenotype) and activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in the cell suspension obtained from a human skin biopsy
The cytoimmunogram (Fig. 37) shows the ratio of viable keratinocytes and their activated forms in the cell suspension obtained from human skin using flow cytometry. The aim was to determine the fraction of viable keratinocytes (CD49f⁺) and, among them, the proportion of activated cells (CD49f⁺ HLA-DR⁺), which may reflect regenerative activity of the epidermis and the degree of immune activation.
X-axis — CD49f-PE.
A marker of stem and mature basal keratinocytes (adhesion integrin α6β1). Increased expression denotes activation of the keratinocyte phenotype.
Y-axis — HLA-DR-PC7.
A marker of immune activation. Not normally expressed by keratinocytes; appears during inflammation, injury, or stimulation.
The total proportion of viable keratinocytes (CD49f⁺) was 95.6% (B2 + B4 = 41.4% + 54.2%), indicating an exceptionally high representation of epidermal cells in the biopsy sample and excellent material quality.
The activation level among viable keratinocytes was 43.3%, meaning that a substantial fraction of cells was in an activated state. This may reflect inflammatory activity, reparative processes, exposure to topical or systemic agents, or in vitro stimulation.
Non-viable cells (debris) accounted for only 3.1%, confirming high overall viability of the suspension and proper cell-isolation technique.
Thus, the high proportion of CD49f⁺ cells confirms that epidermal keratinocytes predominate in the sample, while the elevated percentage of HLA-DR⁺ keratinocytes indicates pronounced functional activity within this population—typical of:
• cutaneous regeneration,
• effects of immunomodulatory agents or irritants,
• early wound-healing phases,
• or inflammatory activation.
In this same skin sample, 86% of fibroblasts were viable, and 6.2% of them were in an activated state (Figs. 38 and 39).
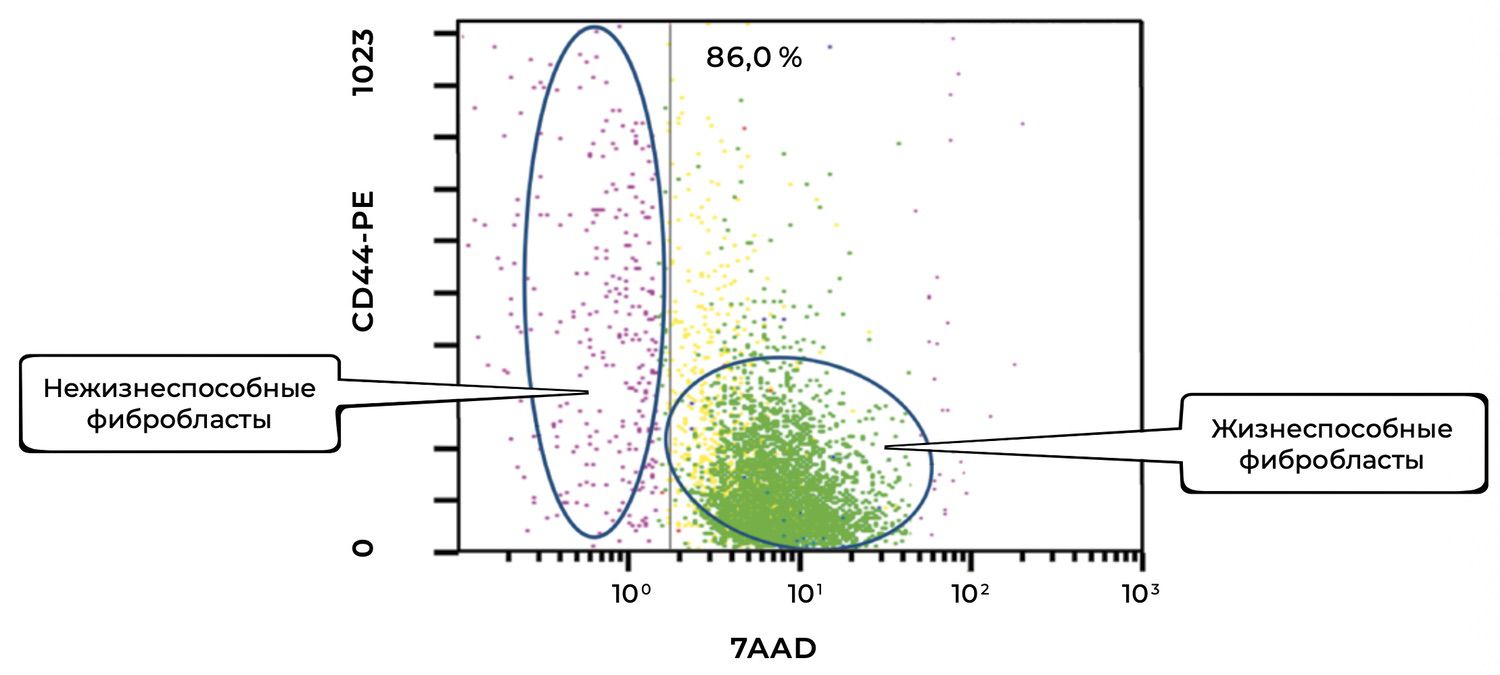
The cytoimmunogram in Figure 38 illustrates the viability of fibroblasts in the cell suspension isolated from human skin. The aim was to determine the proportion of viable dermal fibroblasts (CD44⁺ CD45⁻ CD14⁻) among all acquired events, based on 7AAD expression as the viability marker.
X-axis — 7AAD (viability marker).
An intercalating fluorescent dye that penetrates only cells with compromised membranes:
• Low expression → viable cells
• High expression → non-viable cells
Y-axis — CD44-PE (fibroblast marker).
A surface glycoprotein characteristic of dermal fibroblasts and cells of mesenchymal origin. In combination with exclusion of CD45 and CD14 (lymphocytes/monocytes), it enables precise identification of fibroblast populations.
The high proportion of viable fibroblasts (86%) indicates:
High quality of the isolated sample,
Minimal cytotoxic damage during biopsy processing,
Suitability of the cell suspension for subsequent in vitro / ex vivo procedures (e.g., stimulation assays, culturing, functional profiling).
The CD44⁺ phenotype confirms that the population indeed represents dermal fibroblasts, provided that CD45 and CD14 are excluded.
The small residual proportion of non-viable fibroblasts (left cluster) does not exceed a critical threshold and does not interfere with downstream analyses.
Thus, this cytoimmunogram complements the previous data on keratinocyte viability and allows evaluation of both the epidermal and dermal components of the skin cell suspension. This is especially important when the study aims to assess the regenerative potential of the skin, inflammatory processes, responses to therapeutic agents, or preparation of cell material for culturing.
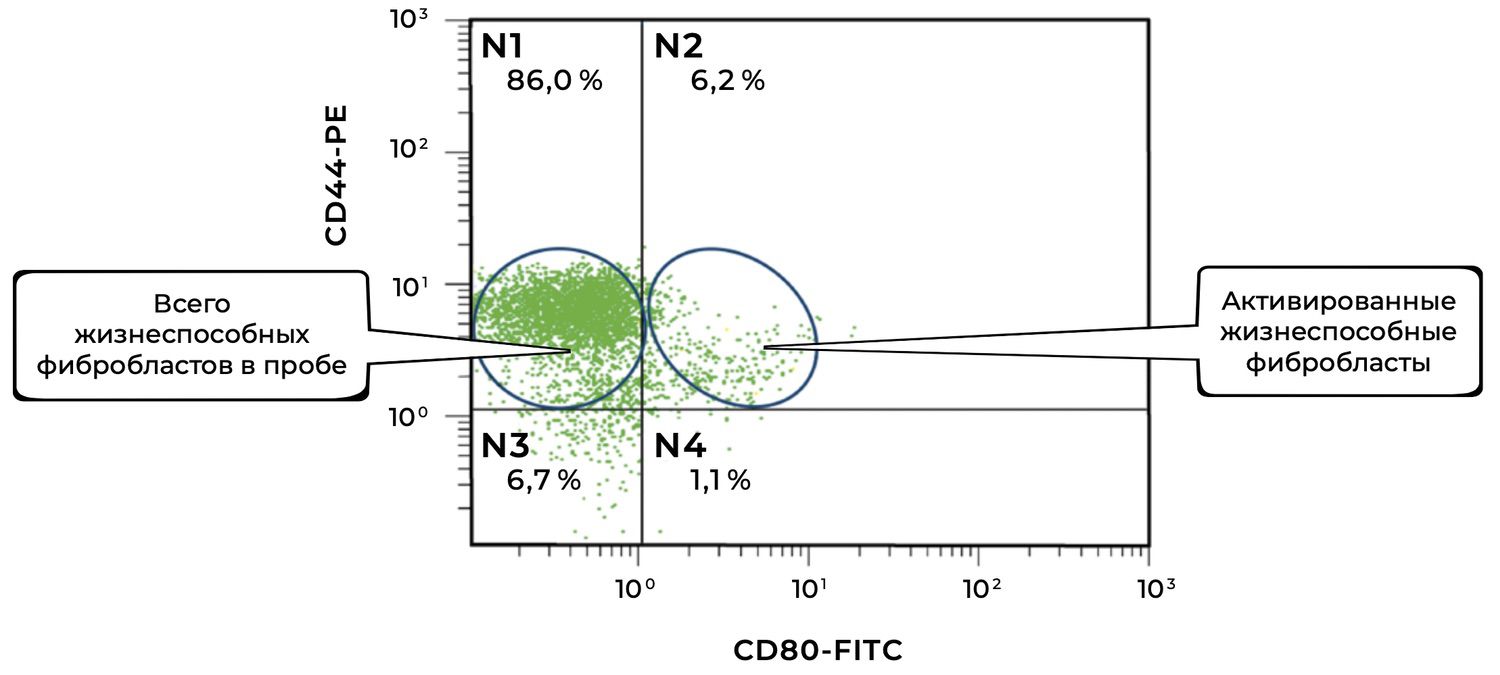
The cytoimmunogram in Figure 39 illustrates the ratio of viable and activated fibroblasts within the cell suspension obtained from a human skin biopsy. This enables determination of the proportion of viable dermal fibroblasts (CD44⁺ CD45⁻ CD14⁻) that exhibit signs of functional activation, i.e., expression of the co-stimulatory molecule CD80.
X-axis — CD80-FITC.
CD80 is a classical activation marker, particularly relevant to antigen presentation and intercellular immune signaling.
• CD80⁻ — non-activated fibroblasts
• CD80⁺ — activated fibroblasts
Y-axis — CD44-PE.
Surface marker of dermal fibroblasts. Together with the exclusion of CD45 and CD14, it enables precise isolation of a pure fibroblast population.
The total proportion of viable fibroblasts in the sample (CD44⁺) is 92.2% (N1 + N2), which confirms the high quality of the cellular material.
The proportion of activated viable fibroblasts (CD44⁺ CD80⁺) is 6.2%, indicating the presence of a local or induced cellular response, suggesting that fibroblasts may be involved in immunomodulation and wound-healing regulation, or reacting to an external stimulus (e.g., inflammation, topical agents, UV exposure).
The majority of fibroblasts are in a non-activated state (86%), which may correspond to the condition of normal skin or post-inflammatory recovery.
CD80⁺ fibroblasts exhibit immunological activity, capable of interacting with both innate and adaptive immune cells. In the context of phenotypic dermatology, this may indicate:
a response to injury or stress,
the onset of tissue remodeling,
participation of fibroblasts in antigen presentation.
Thus, the cytoimmunogram complements the previously presented data on overall cell viability and shows that, despite the high proportion of viable fibroblasts, only a small fraction is activated. This may be important for assessing therapeutic efficacy, inflammatory status, and the regenerative potential of the dermis.
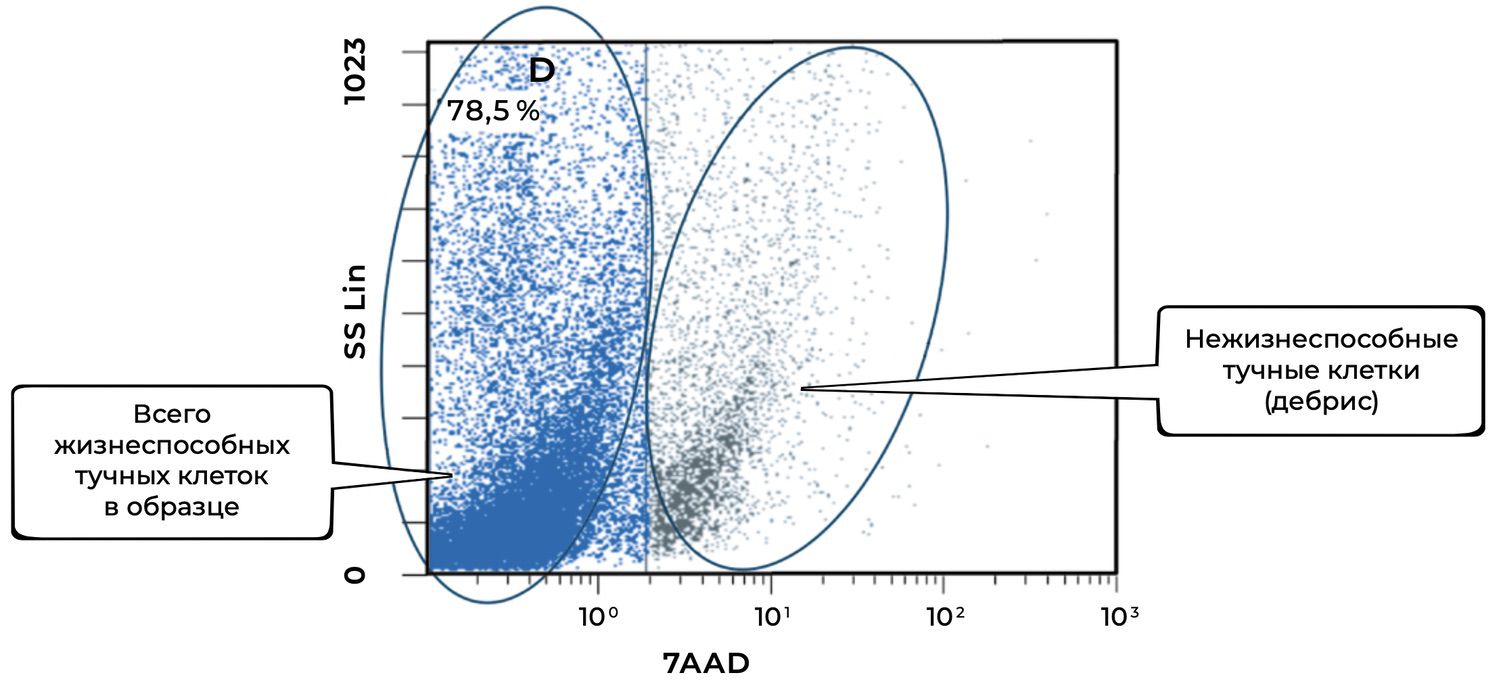
The cytoimmunogram in Figure 40 illustrates the assessment of mast-cell viability in the cell suspension derived from the skin biopsy. It allows determination of the proportion of viable mast cells (CD249⁺, also known as c-Kit/SCFR) in the total cellular pool of the biopsy using the viability marker 7AAD, which identifies non-viable cells.
X-axis — 7AAD.
A marker of apoptosis and cell death:
• 7AAD⁻ — viable cells
• 7AAD⁺ — non-viable cells (debris, fragmented cells)
Y-axis — SS Lin (side scatter).
Reflects granularity and internal complexity; mast cells typically exhibit high side-scatter due to their granules.
The total proportion of viable mast cells in the sample is 78.5%, which indicates:
good sample quality,
appropriate conditions of biopsy collection and storage,
high functional suitability of the material for downstream analysis (e.g., evaluation of degranulation, activation markers, etc.).
The remaining ~21.5% of cells are non-viable, likely due to degradation, mechanical damage, or late-stage activation (autodestruction).
Mast cells are key mediators of the cutaneous immune response: they participate in early phases of inflammation, contain histamine, tryptase, and chemokines, and play essential roles in angiogenesis and wound healing. A high proportion of viable mast cells confirms that the sample:
contains a preserved pool of functional mast cells,
allows further activation studies and phenotyping (e.g., CD107a, additional degranulation markers),
can be used to assess their involvement in cutaneous immunopathogenesis or wound-healing processes.
Thus, this cytoimmunogram completes the comprehensive phenotypic evaluation of the cellular composition of the skin biopsy. Together with fibroblasts and keratinocytes, viable mast cells constitute another important component potentially involved in inflammatory, allergic, or regenerative processes of the skin.
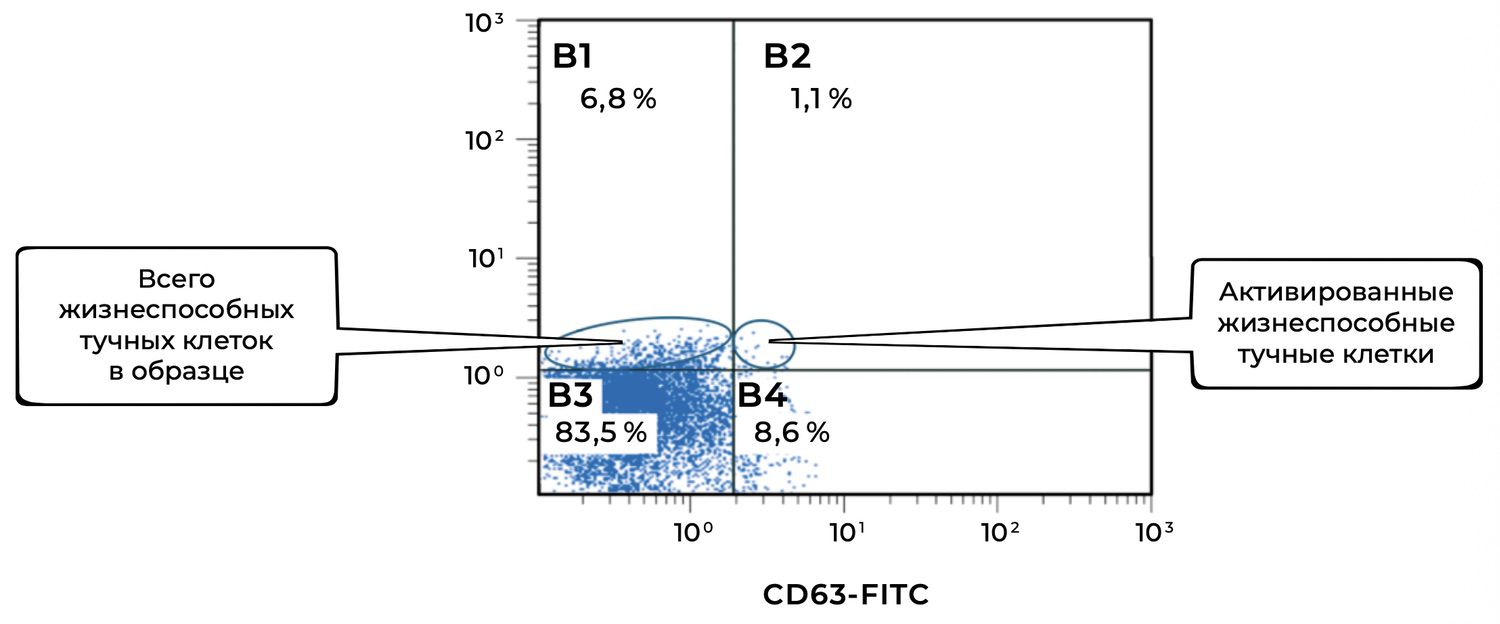
The cytoimmunogram illustrating the ratio of viable and activated mast cells in the biopsy-derived cell suspension (Fig. 41) identifies, among viable mast cells (CD249⁺), the proportion that exhibits signs of activation (CD63⁺). This analysis allows assessment of the level of local inflammatory activity and mast-cell degranulation.
X-axis — CD63-FITC.
A marker of mast-cell activation; CD63 is expressed on the cell surface during degranulation.
Y-axis — CD249-PE (c-Kit, SCFR).
A specific marker of mast cells, defining their identity within the population.
The total proportion of viable mast cells in the sample is 83.5% + 8.6% = 92.1%, which confirms the preservation and integrity of the mast-cell population within the skin biopsy.
Among them, 8.6% are activated, which may indicate:
a local inflammatory reaction (e.g., in areas of chronic dermatitis, psoriasis, allergy),
background or stimulus-induced mast-cell degranulation.
CD63 is used as a classical indicator of degranulation — the release of histamine and pro-inflammatory mediators. An increased proportion of CD63⁺ cells may be clinically relevant for:
evaluating the effectiveness of therapy (e.g., mast-cell membrane stabilizers),
diagnosing hypersensitivity reactions.
CD249⁺ CD63⁺ mast cells represent a clear marker of mast-cell activation and participation in inflammatory processes.
A decrease in this population may indicate effective antihistamine or anti-inflammatory therapy.
An increase may reflect exacerbation or an active stage of a dermatological disease (such as atopic dermatitis, contact eczema, rosacea, etc.).
Thus, in this skin sample, viable mast cells constitute the majority of the population, with approximately one-tenth in an activated state. This makes the diagram highly informative for assessing the cutaneous immune status, monitoring therapy, and performing phenotypic dermatological diagnostics.
At the same time, the same sample shows 7% monocytes, of which 2.6% are in an activated state (Fig. 42).
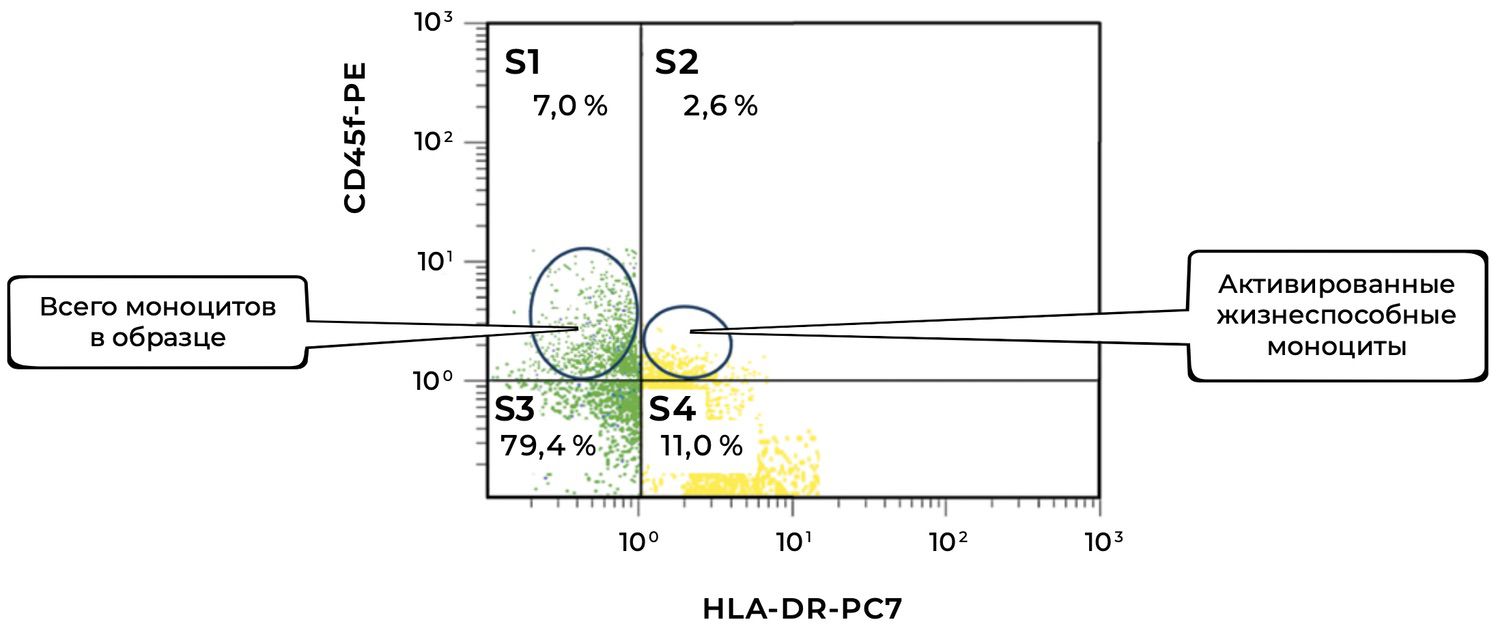
The cytoimmunogram (Fig. 42) illustrates the ratio of viable and activated monocytes in the skin-biopsy cell suspension and allows determination of the proportion of viable monocytes (CD45⁺ CD14⁺) that are in an activated state, i.e., expressing HLA-DR — an MHC class II molecule involved in antigen presentation.
X-axis — HLA-DR-PC7.
A marker of monocyte activation and antigen-presenting cells.
Y-axis — CD45-PE.
A pan-leukocyte marker used to identify immune cells, including monocytes.
The total proportion of viable monocytes in the sample is 79.4% + 11.0% = 90.4%, which represents an excellent level of cellular preservation.
Among them, 11.0% express HLA-DR, which may indicate:
involvement in a local inflammatory process,
functional activity characteristic of antigen-presenting cells,
the presence of immune activation within the dermis (e.g., in autoimmune dermatoses, microbial inflammation, tissue injury, etc.).
CD45⁺ CD14⁺ HLA-DR⁺ monocytes are an important marker of the innate immune response.
An elevated proportion in a skin biopsy may suggest:
enhanced antigen presentation,
immune infiltration during chronic inflammation.
Conversely, a decreased proportion may indicate immunosuppression or poor tissue representativeness.
In this skin sample, the majority of monocytes are viable, enabling functional and phenotypic analysis, and approximately 11% are activated, which may reflect the involvement of innate immunity in the pathogenesis of the local process. This is highly relevant for phenotypic dermatology, particularly in the evaluation of inflammatory and autoimmune skin conditions.
Viable intraepidermal macrophages constitute 47.9% of this skin sample, with 4.9% of them in an activated state (Fig. 43).
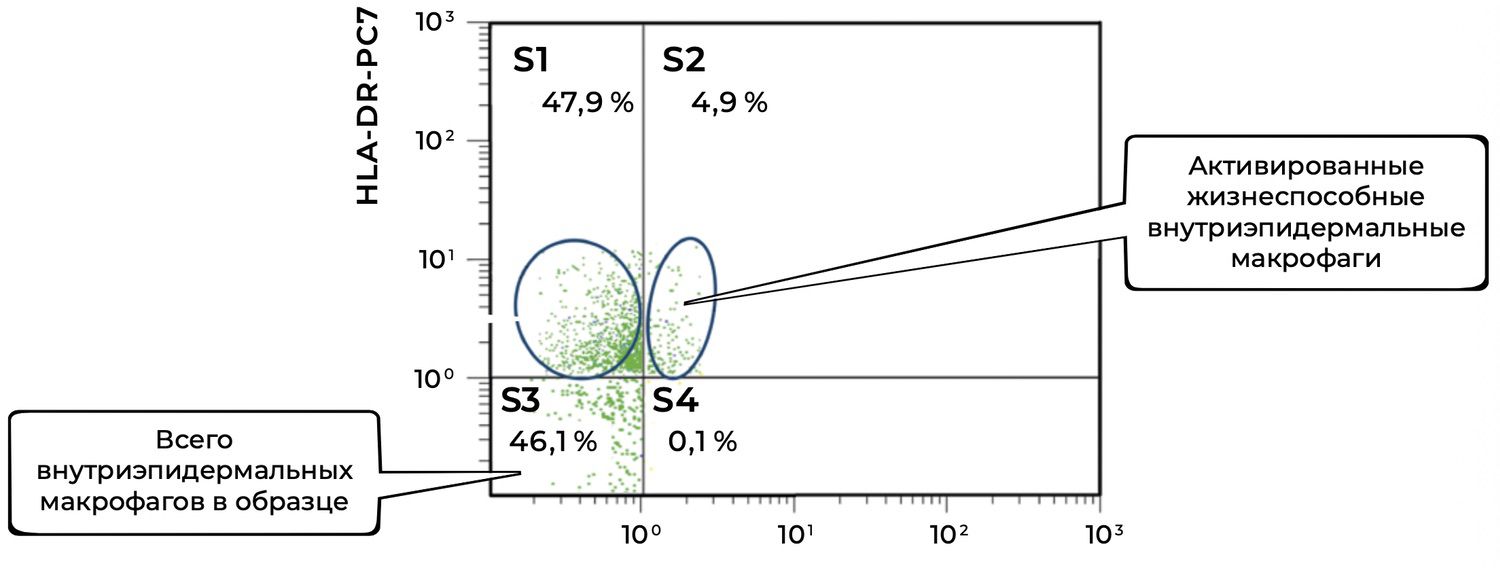
The cytoimmunogram (Fig. 43) illustrates the ratio of viable and activated intraepidermal macrophages in the cell suspension derived from the skin biopsy. It quantifies:
the proportion of viable intraepidermal macrophages (identified by CD207 / langerin expression), and
the fraction among them that is activated (HLA-DR⁺).
X-axis — CD207-PE.
A marker of intraepidermal macrophages (Langerhans cells), specific to the epidermis.
Y-axis — HLA-DR-PC7.
A marker of functional activation characteristic of antigen-presenting cells.
The total number of CD207⁺ cells (intraepidermal macrophages) is:
S3 + S4 = 46.1% + 0.1% = 46.2%.
Among them, only 0.1% express HLA-DR, which may indicate:
a resting or tolerant state;
a low level of immune stimulation at the time of biopsy;
a physiologic absence of activation in normal skin.
The high proportion of S1 (47.9%) may include other dermal antigen-presenting cells as well as background events requiring additional evaluation.
CD207⁺ HLA-DR⁺ cells represent functionally active Langerhans cells, capable of migration and antigen presentation to T cells. Their extremely low proportion (0.1%) may be characteristic of:
normal, uninfected skin,
a physiological state of homeostasis,
or, conversely, may reflect depletion or deactivation in chronic inflammation.
Thus, in this patient’s skin biopsy, there is a high overall representation of CD207⁺ intraepidermal macrophages, but virtually no activation (as indicated by HLA-DR expression). This may be normal in the absence of a stimulus, but must also be interpreted in the broader context of the skin’s immune profile — including monocytes, mast cells, fibroblasts, and keratinocytes.
In this same skin sample, endothelial cells are represented by 5.4% singly activated and 5.9% doubly activated, with a total of 60.3% viable endothelial cells (Fig. 44).
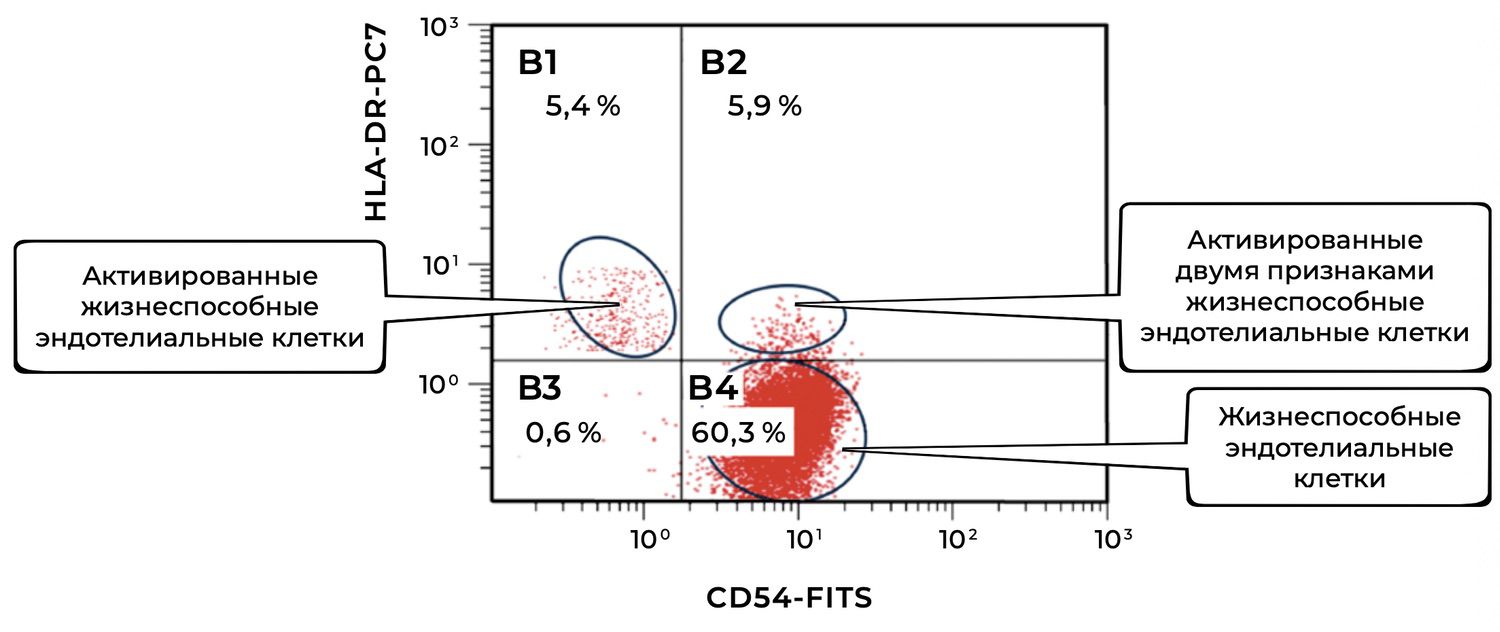
The cytoimmunogram (Fig. 44) reflects the ratio of viable and activated endothelial cells (CD146⁺ phenotype) in the cell suspension obtained from the skin biopsy. It allows determination of:
the relative number of viable endothelial cells (CD146⁺), and
the identification of activated forms based on one or two additional activation markers:
HLA-DR⁺ — a marker of antigen presentation;
CD54⁺ (ICAM-1) — a marker of inflammatory activation and adhesion;
CD54⁺ HLA-DR⁺ — combined activation.
The parameters of this cytoimmunogram are as follows:
X-axis — CD54-FITC.
A surface adhesion molecule (ICAM-1), upregulated during inflammation.
Y-axis — HLA-DR-PC7.
A class II MHC molecule — a marker of antigen presentation, activation, and immune activity.
The total proportion of viable endothelial cells (CD146⁺) in the sample is:
B4 + B2 + B1 + B3 = 60.3% + 5.9% + 5.4% + 0.6% = 72.2%.
Among these, the activated subsets include:
HLA-DR⁺ only: 5.4%
CD54⁺ only: 5.9%
Dual activation (CD54⁺ HLA-DR⁺): 0.6%
Total activated forms: 11.9% (summed without overlap because the diagram logically separates the subpopulations)
Physiological significance
CD146⁺ cells represent endothelial cells of the dermal microvasculature, involved in the regulation of vascular tone, permeability, and leukocyte migration.
Activated forms (CD54⁺ and/or HLA-DR⁺) reflect:
local inflammatory signaling,
participation in immune responses,
or endothelial activation in response to tissue injury.
The proportion of activated endothelial cells (~12%) can vary physiologically but typically reflects a moderate level of baseline immune tone in the skin.
Thus, in this skin biopsy, non-activated viable endothelial cells dominate (60.3%), while approximately 12% exhibit immune activation, indicating potential involvement of the microvascular endothelium in low-grade immunologic surveillance or post-inflammatory remodeling.
The results obtained via flow cytometry as a demonstration of precision diagnostics in skin immunophenotyping may be interpreted as follows:
There is a robust representation of keratinocytes, with the majority being activated, which indicates proliferative activity of the basal epidermal layer.
B lymphocytes are present; although they normally reside in circulating blood and lymph, their presence in skin suggests humoral immune activity.
Several subtypes of T lymphocytes (CD3⁺) are present, localized primarily in the outer three layers of the epidermis.
The fact that CD4⁺ cells slightly outnumber CD8⁺ cells indicates enhancement of the adaptive immune component.
Low levels of T suppressor cells suggest an absence of infectious or acute inflammatory processes.
The remaining indices show modest numbers of specialized skin-resident cells with low activation levels, which—combined with the absence of clinical symptoms—indicate a normal physiological state of the skin in this individual.

These conclusions are based on direct analysis of the cellular composition, viability, and activation state of the populations in a single skin biopsy sample, as assessed by high-parameter flow cytometry.
This approach allows simultaneous evaluation of overall viability and activation of key cellular populations involved in maintaining skin homeostasis and regulating inflammatory responses.
To simplify interpretation and better demonstrate diagnostic value, the proportions of viable and activated cells for each population are summarized below.
Viable cells: 54.2%
Activated cells (CD49f⁺ HLA-DR⁺): 41.4%
Comment: A large proportion is activated, suggesting immune or reparative activity in the epidermis.
Viable cells: 86.0%
Activated cells (CD44⁺ CD80⁺): 6.2%
Comment: High viability with low activation is typical of resting dermal fibroblasts.
Viable cells: 83.5%
Activated cells (CD249⁺ CD63⁺): 8.6%
Comment: Mast-cell activation may indicate involvement in allergic or inflammatory reactions.
Viable cells: 79.4%
Activated cells (CD45⁺ CD14⁺ HLA-DR⁺): 11.0%
Comment: The presence of activated monocytes points to local immune activity and potential antigen-presentation dynamics.
Viable cells: 47.9%
Activated cells (CD207⁺ HLA-DR⁺): 4.9%
Comment: Lowest viability among cell types, likely due to fragility during enzymatic and mechanical isolation.
Viable cells: 60.3%
Activated forms:
CD146⁺ CD54⁺: 5.9%
CD146⁺ HLA-DR⁺: 5.4%
CD146⁺ CD54⁺ HLA-DR⁺: included within both of the above
Comment: Expression of ICAM-1 (CD54) and HLA-DR suggests participation of the vascular endothelium in inflammatory signaling and leukocyte recruitment.
Thus:
Fibroblasts and mast cells demonstrated the highest viability, consistent with their intrinsic resilience and stable presence within the dermis.
The greatest degree of activation was observed in keratinocytes (likely in response to injury or inflammation), as well as in monocytes and endothelial cells.
The lowest viability was recorded in intraepidermal macrophages, which may reflect their high sensitivity to mechanical and enzymatic isolation.
A differentiated assessment of activation status—using multiple markers—provides a more accurate understanding of the functional state of each cell population.
This example demonstrates the applicability of phenotypic analysis in a broader cohort of volunteers to determine sex- and age-related characteristics of the quantitative and functional state of skin cells.
In 80 healthy volunteers—divided into sex- and age-stratified groups of 16 individuals each—skin punch biopsies were performed. Subpopulations of cells were isolated from each biopsy specimen for immunophenotyping and for constructing individual skin cytoimmunograms.
Statistical analysis was carried out using descriptive methods, including arithmetic means and their standard errors (M ± m), with application of the Student’s t-test to determine the statistical significance of differences between mean values, assuming a normal distribution of the source data.⁵⁷
As a result, from the total heterogeneous population of skin cells we obtained distinct viable subpopulations from both native and cryopreserved samples. For each, the cellular phenotype, functional activity, and viability were determined (Table 1).
Table 1. Comparative assessment of the quantitative and functional state of viable cells in native and cryopreserved skin biopsy samples from conditionally healthy individuals, n = 80
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Нативный образец, % |
Криоконсервированный образец, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
70,25±3,75 3,25±0,75 |
68,2±2,01 1,3±0,04 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
76,5±3,5 4,93±2,47 |
66,8±4,0 3,5±0,2 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
48±1,0 3,8±0,9 5,1±1,1 3,7±0,7 |
46,4±1,2 3,6±0,8 1,3±0,05 3,5±0,9 |
|
CD146+ CD146+ CD 54– HLA-DR+ CD146+ CD 54+ HLA-DR– CD146+ CD 54+ HLA-DR+ CD146+ CD 34+ |
1,32±0,98 0 22,88±2,52 0,33±0,17 6,93±1,07 |
0,6±0,03 0 50,0±4,6 0,40±0,12 36,0±5,2 |
|
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
3,43±1,77 1,1±0,2 |
2,5±0,99 1,6±0,3 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
7,75±1,25 0,23±0,16 |
5,6±1,01 0,28±0,14 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
14,0±1,0 10,5±1,5 2,5±0,5 6,0±1,0 10,5±1,5 |
11,2±1,99 9,9±0,75 1,3±0,22 7,0±1,05 9,5±1,14 |
|
Жизнеспособность, % |
99,8±0,9 |
87,0±0,5 |
When comparing the relative proportions of skin cell populations between native and cryopreserved samples, the following results were obtained.
Overall cell viability after cryopreservation remained high — 87.0 ± 0.5%, which corresponds to approximately 87% of the native value (99.8 ± 0.9%). This indicates preservation of cellular structures and membrane integrity under the applied cryoprotective conditions.
Keratinocytes (CD49f⁺ HLA-DR⁺) and fibroblasts (CD45⁻ CD14⁻ CD44⁺) demonstrated a slight decrease in cell counts in cryopreserved samples (~10%), while the proportion of activated forms remained within the range of statistical variability. This suggests preservation of the basal metabolic and reparative potential of the tissue.
Langerhans cells (CD207⁺ CD80⁺ HLA-DR⁺) and their activated subpopulations showed a moderate decrease in activity (by ~3–5%) after freezing; however, the DR⁺/DR⁻ ratio remained stable. This indicates preservation of epidermal antigen-presenting capacity.
More pronounced changes were observed in the group of endothelial cells (CD146⁺) and their subtypes — the proportion of HLA-DR⁺ cells decreased almost twofold (from 6.9% to 3.6%), reflecting higher sensitivity of vascular endothelium to cryogenic stress.
Mast cells (CD249⁺ CD63⁺) and monocytes (CD45⁺ CD14⁺) showed a moderate reduction in the proportion of activated cells, while total cell counts remained stable, indicating relative resistance of these subpopulations to freezing.
Among epidermal lymphocytes, a decrease was observed in the total proportion of T-cells (CD45⁺ CD3⁺) and T-helpers (CD4⁺), accompanied by preservation or slight increase of cytotoxic CD8⁺ cells. This reflects a shift in the phenotypic balance toward regulatory–suppressor components after cryopreservation.
The cryopreservation protocol ensures overall cell viability above 85% with minimal changes in subpopulation composition.
The most cryo-resistant subpopulations were fibroblasts, keratinocytes, and mast cells, while endothelial and antigen-presenting cells proved more sensitive.
The developed protocol allows reliable subsequent phenotypic analysis and the construction of valid skin cytoimmunograms in both native and cryopreserved samples.
This result supports the use of cryopreservation as a method for preserving samples for longitudinal (dynamic) observation. Stratification of participants by sex and age was necessary to demonstrate differences in measurable parameters among distinct demographic groups.
The resulting mean statistical characteristics (Table 2, Parts 1 and 2) are provided to demonstrate the potential of screening approaches for assessing the phenotypes of skin cell subpopulations.
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Мужчины, n=40 |
||||
|
15–25 лет |
26–35 лет |
36–45 лет |
46–55 лет |
56–65 лет |
||
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
74,98±2,11* 4,98±0,32* |
70,25±3,75 3,25±0,75 |
69,5±3,5 3,23±0,37 |
61,0±5,0 2,78±0,62 |
55,75±2,25* 2,25±0,65* |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
82,75±1,25* 7,15±1,15* |
76,5±3,5 4,93±2,47 |
72±5,0 4,13±1,27 |
66,5±3,5 2,4±0,47 |
61,25±1,75* 3,18±0,82* |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
52,0±4,0* 0,18±0,02* 5,48±0,62 0,25±0,05 |
48±1,0 3,8±0,9 5,1±1,1 0 |
45,5±4,5 0,27±0,13 5,25±0,95 0,73±0,27 |
43,0±2,0 1,3±0,4 5,7±1,3 0,3±0,1 |
36,0±3,0* 1,15±0,35* 7,5±1,5 0 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
0,9±0,1 0 27,38±1,62* 4,15±0,85* 11,52±2,62* |
1,32±0,98 0 22,88±2,52 0,33±0,17 6,93±1,07 |
1,03±0,17 0,18±0,02 22,23±0,17 1,35±0,25 3,35±0,27 |
1,28±0,12 0 19,73±0,17* 1,43±0,27 1,42±0,08 |
1,1±0,2 0,08±0,02 18,85±1,55* 1,32±0,42* 0,43±0,07* |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
6,0±0,2* 3,63±1,27* |
3,43±1,77 1,1±0,2 |
4,23±0,77 2,4±0,3 |
3,78±0,42 1,15±0,05 |
4,06±0,43* 0,6±0,1* |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
6,8±0,4* 2,45±0,15* |
7,75±1,25 0,23±0,16 |
6,9±0,3 2,15±0,45 |
5,83±1,17 1,28±0,4 |
4,58±0,42* 0,3±0,02* |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
15,75±1,25 11,5±2,5 3,75±1,75 7,5±0,5 9,75±2,25 |
14,0±1,0 11,0±1,0 2,5±0,5 6,0±1,0 10,5±1,5 |
13,25±0,75 7,23±3,77 4,0±1,0 6,25±1,75 10,75±1,25 |
13,75±1,25 10,5±0,5 3,25±1,75 5,25±1,75 7,5±0,5 |
12,5±1,5 9,75±0,25 2,75±1,25 4,5±1,5 6,5±1,5 |
|
Жизнеспособность, % |
84,75±4,25 |
86,0±4,0 |
88,5±1,5 |
84,75±4,25 |
83,25±5,75 |
|
In the first part of Table 2, the results of flow-cytometric analysis of skin-derived cell suspensions are presented. These data reflect the quantitative distribution and phenotypic activity of the main subpopulations of skin cells in different male age groups (15–65 years). The indicators are expressed as percentages of the total number of viable cells (mean ± standard deviation).
The main observations are summarized below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
A consistent age-related decline in the proportion of viable and activated keratinocytes is observed: from 74.98 ± 2.11% in the 15–25 year group to 55.75 ± 2.25% in men aged 56–65 years (p<0.05). This reflects age-related depletion of the epidermal regenerative potential.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺).
A similar pattern is seen: a gradual decrease in the number of fibroblasts and their activated forms (CD80⁺) after the age of 35 (from 82.75 ± 1.25% to 61.25 ± 1.75%), indicating reduced dermal matrix metabolism and reparative activity.
Langerhans cells (CD207⁺).
Both the number and activation level of Langerhans cells decrease after the age of 35, most prominently after 55 years (from 52.0 ± 4.0% to 36.0 ± 3.0%, p<0.05). This may indicate a decline in epidermal antigen-presenting function and age-associated immune remodeling.
Endothelial cells (CD146⁺).
Age-related decreases occur predominantly among DR⁺ subtypes (from 11.5 ± 2.6% to 0.43 ± 0.07%), reflecting a reduction in angiogenic activity and slower neovascular processes in aging skin.
Mast cells (CD249⁺ CD63⁺).
Overall values remain relatively stable across age groups (3.4–4.0%), but activated forms (CD63⁺) show a slight increase after age 45, possibly indicating a rise in baseline inflammatory activity.
Monocytes (CD45⁺ CD14⁺).
The proportion of activated monocytes (HLA-DR⁺) decreases in older groups (from 2.45 ± 0.15% to 0.3 ± 0.02%), suggesting a shift from active inflammatory response toward a more regulatory immune profile.
Epidermal lymphocytes (CD45⁺ CD3⁺).
A general reduction in the total T-cell pool and in T-helper cells (CD4⁺) is observed with age, while cytotoxic T cells (CD8⁺) remain preserved. This indicates an age-related decline in the adaptive immune component with relative maintenance of cytotoxic potential.
Cell viability.
Overall skin-cell viability declines from 88.5 ± 1.5% in men aged 15–25 years to 83.25 ± 5.75% in the 56–65 year group, confirming the general age-related reduction of reparative and immune reserves in the skin.
Conditionally healthy male skin demonstrates a clear age-related phenotypic dynamic characterized by:
a decline in regenerative and immunocompetent cell populations,
reduced activity of antigen-presenting and endothelial components,
a tendency toward predominance of suppressive and low-reactivity cellular phenotypes.
These findings confirm that cellular and phenotypic age of the skin may serve as a more accurate indicator of its condition than chronological age and highlight the necessity of applying phenotypic dermatology for patient stratification and prediction of therapeutic responses.
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Женщины, n=40 |
||||
|
15–25 лет |
26–35 лет |
36–45 лет |
46–55 лет |
56–65 лет |
||
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
73,25±1,75* 3,08±0,92 |
66,5±2,5 3,93±0,07 |
67,25±1,75 4,3±0,9 |
55,66±3,33* 1,97±0,23 |
43,33±3,67* 2,05±0,45 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
78,5±0,5* 9,55±0,85* |
71,25±0,75 4,68±0,72 |
71,25±2,75 5,4±1,0 |
61,0±1,0* 2,0±0,1* |
55,0±2,0* 0,4±0,2* |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
46,5±1,5* 1,95±0,95 7,6±0,8 0,25±0,15 |
46,5±0,5 0,15±0,05 6,53±2,17 0,55±0,25 |
45,25±0,75 2,13±0,77 6,22±2,08 0,63±0,27 |
42,25±1,75 1,63±0,37 7,58±0,72 0,4±0,1 |
37,5±1,5* 0,9±0,01 8,65±1,35 0,45±0,05 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
1,05±0,65 0,23±0,07 26,95±1,75* 0 11,35±2,05* |
0,75±0,15 0 24,5±0,6 1,03±0,17 5,1±1,0* |
0,95±0,15 0 21,28±1,92 0,36±0,14 3,83±0,47* |
0,92±0,08 0,4±0,1 19,25±1,75* 1,1±0,1 1,67±0,63* |
0,95±0,2 0,75±0,25 16,75±0,25* 0 0,17±0,03* |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
4,53±0,67 0,85±0,15 |
2,4±0,5* 0,68±0,3 |
3,2±1,0 1,1±0,5 |
3,72±0,28 1,38±0,12 |
4,48±0,92 0,96±0,53 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
10,4±3,6* 0,67±0,19 |
7,03±0,03 1,13±0,37 |
8,5±1,5 1,95±0,45 |
4,5±0,3* 0,9±0,2 |
4,5±1,4* 0,26±0,06 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
15,0±0,1 10,5±1,5* 4,5±0,5 7,0±1,0* 9,0±1,0 |
14,25±0,75 10,25±1,75 4,0±2,0 7,0±1,0 9,0±1,0 |
13,0±2,0 9,5±1,5 3,5±0,5 7,0±0,01 9,25±0,75 |
11,5±0,5 8,25±1,75 3,0±1,0 4,75±1,25 8,75±1,25 |
11,0±2,0 5,3±1,7* 3,75±1,25 4,5±0,5* 8,5±1,5 |
|
Жизнеспособность, % |
89,25±3,75 |
90,25±1,75 |
85,25±1,75 |
89,5±1,5 |
89,5±2,5 |
|
In the second part of Table 2, the results of phenotypic analysis of the skin-cell composition in women aged 15 to 65 years are presented, obtained using flow cytometry.
The table shows the mean percentages of major cell subpopulations and their activated forms, characterizing the morphofunctional state of the epidermis and dermis across different age groups.
The main observations and trends are summarized below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
Women demonstrate a clear age-dependent decline in the proportion of viable and activated keratinocytes — from 73.25 ± 1.75% in the 15–25 year group to 43.33 ± 3.67% in the 56–65 year group (p<0.05). This reflects decreased epidermal renewal rates and weakening of barrier–regenerative processes after age 45.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺).
The proportion of fibroblasts decreases by nearly 30% in older age groups (from 78.5 ± 0.5% to 55.0 ± 0.6%), while activated forms (CD80⁺) show a particularly marked decline after age 50. This indicates age-related reduction in dermal synthetic activity and diminished extracellular-matrix remodeling capacity.
Langerhans cells (CD207⁺ CD80⁺ HLA-DR⁺).
Their numbers gradually decrease from 46.5 ± 1.5% in younger women to 37.5 ± 1.5% after age 55, while activated forms (CD80⁺ HLA-DR⁺) decline nearly twofold. This reflects reduced antigen-presenting capacity and diminished local immune surveillance in the skin.
Endothelial cells (CD146⁺ CD54⁺ CD34⁺).
A pronounced age-related decrease is observed in activated phenotypes (CD54⁺ CD34⁺), indicating reduced angiogenic activity and microcirculatory potential of the skin. The most significant differences are seen between the 15–25 and 56–65 age groups (p<0.05).
Mast cells (CD249⁺ CD63⁺).
Their overall proportion remains stable (around 3–4%), but the number of activated cells (CD63⁺) slightly increases after age 45. This reflects a trend toward enhanced sensitization and mast-cell involvement in age-associated inflammatory reactions.
Monocytes (CD45⁺ CD14⁺ HLA-DR⁺).
With age, their total number declines by half, and activated forms (HLA-DR⁺) fall more than threefold, indicating fading inflammatory activity and a shift toward a more regulatory immune profile.
Epidermal lymphocytes (CD45⁺ CD3⁺).
Women also show an age-related decrease in total T-lymphocyte counts, particularly T-helpers (CD4⁺), while cytotoxic cells (CD8⁺) and B-lymphocytes (CD19⁺) remain relatively stable. This reflects a general weakening of the adaptive immune response while maintaining baseline cytotoxic capacity.
Skin-cell viability.
Viability remains high in women and varies only slightly (85–90%), indicating greater resilience of female skin cells to age-related changes compared with men.
Female skin demonstrates a more stable phenotypic profile than male skin, yet it also shows a gradual decline in regenerative and immune activity with age. The most pronounced changes occur in the dermal (fibroblast) and antigen-presenting (Langerhans cell) compartments, while epidermal and mast-cell populations remain relatively stable. Thus, these findings highlight sex-specific features of skin aging and confirm the value of the phenotypic approach as a tool for identifying cellular signatures of age-related remodeling and for predicting the effectiveness of rejuvenation and restorative therapies.
Comparison of male and female phenotypic profiles reveals pronounced sex differences in the pattern and rate of age-related changes, despite shared general trends in the degradation of regenerative and immune functions.
Across both sexes, aging is characterized by:
gradual reduction of the viable skin-cell pool (by ~10–15% by age 60),
decreased numbers of activated keratinocytes, fibroblasts, and Langerhans cells,
weakening of the antigen-presenting compartment (HLA-DR⁺ phenotypes),
reduced pools of activated monocytes and endothelial cells,
relative stability of mast cells and cytotoxic lymphocytes (CD8⁺ CD56⁺).
These processes represent a universal phenotypic signature of skin aging — a shift from a regenerative–inflammatory state to a metabolically and immunologically hyporeactive one.
Changes develop gradually and linearly starting from age 30–35.
Early and more pronounced declines occur in epidermal and dermal compartments:
keratinocytes decrease by nearly 25% by age 45,
fibroblasts decrease by 20–25% compared to young groups.
A sharper reduction in CD34⁺ regenerative cells and DR⁺ endothelial cells indicates accelerated vascular–regenerative aging.
By age 55–60, male skin exhibits a hypoinflammatory phenotype with depletion of T-helpers and weakened dermal repair.
Decline in cellular activity begins later — after age 45 — but then becomes steeper and more multi-component.
Women maintain higher cell viability (up to 90% in older groups vs. 83% in men), reflecting better physiological tissue resilience.
In postmenopausal age, a pronounced decrease in fibroblasts and Langerhans cells (–30–35%) occurs, reflecting hormone-driven acceleration of dermal and immune aging.
Unlike men, women maintain relatively stable levels of CD8⁺ and NK cells, preserving cytotoxic and antiviral skin activity even at advanced age.
Male skin ages via early regenerative depletion, whereas female skin ages via later, hormonally modulated structural decline.
In both cases, a phenotypic shift occurs—from the dominance of regenerative and immunologically active subpopulations toward the predominance of low-reactive and more differentiated cells.
These differences underscore the need for personalized phenotypic stratification of patients when assessing skin conditions, forecasting aging trajectories, and selecting therapeutic protocols—one of the key goals of phenotypic dermatology.
The examination of conditionally healthy individuals made it possible to establish average reference values for the phenotypic parameters of the skin. The results presented in Table 2 will require further refinement, but the foundation has been laid.
Patient L., a 26-year-old woman, was admitted to the dermatology department of the Tyumen Regional Dermatovenerologic Dispensary with complaints of severe pruritus (including nocturnal itching), pronounced skin dryness and scaling, cutaneous edema, excoriations, and fissures.
According to the patient, the disease had been present for more than 15 years, beginning during puberty with the appearance of lesions on the flexural surfaces of the elbows and popliteal fossae. Atopic dermatitis was diagnosed at the first clinical visit. Exacerbations occurred up to three times annually and consistently required hospitalization. Previous treatments included topical glucocorticosteroids, emollients, antihistamines, and topical calcineurin inhibitors. During the preceding two years, the disease had relapsed continuously, progressing to erythroderma and secondary infection of excoriated lesions. The patient had received intravenous methylprednisolone, courses of antibacterial therapy, and intravenous laser blood irradiation, all producing only temporary improvement. Her hereditary and allergy history was unremarkable, although allergologic testing revealed sensitization to household allergens.
Upon admission, the patient’s condition was classified as severe. She exhibited an asthenic body habitus. Physical examination revealed vesicular breath sounds without wheezing, a respiratory rate of 16 breaths per minute, regular heart sounds with a heart rate of 76 beats per minute, and blood pressure of 110/70 mmHg. The tongue was moist and pink. The abdomen was soft and non-tender. The liver and spleen were not enlarged. Bowel habits were normal, and urinalysis revealed no abnormalities. Thyroid examination was unremarkable. Computed tomography of the lungs demonstrated no abnormalities. Ultrasonography of the abdominal organs and thyroid gland revealed no pathology. Laboratory findings included hemoglobin of 107 g/L and an erythrocyte sedimentation rate of 28 mm/h. Urinalysis, serum biochemistry, and thyroid hormone levels were within reference ranges. Testing for helminthic infection was negative. Serum immunoglobulins A, M, and G were normal. C-reactive protein was 9 mg/L. Total serum IgE was markedly elevated at 5,500 IU/mL.
The pathological process was widespread and characterized by confluent bright erythema involving the trunk and extremities, reaching the clinical stage of erythroderma. The condition was accompanied by cutaneous edema, fissures in the antecubital and popliteal regions, extensive fine lamellar scaling, and numerous linear excoriations distributed over the entire skin surface, including areas inaccessible to the patient’s hands. Fissuring and scaling of the vermilion border of the lips (cheilitis) were also present. No visual signs of pyoderma were observed. The SCORAD index was 82. Bilateral inguinal lymph nodes were enlarged to approximately the size of a hazelnut; they were mobile, painless, and soft-elastic on palpation. Other lymph node groups were unaffected (Figure 31).
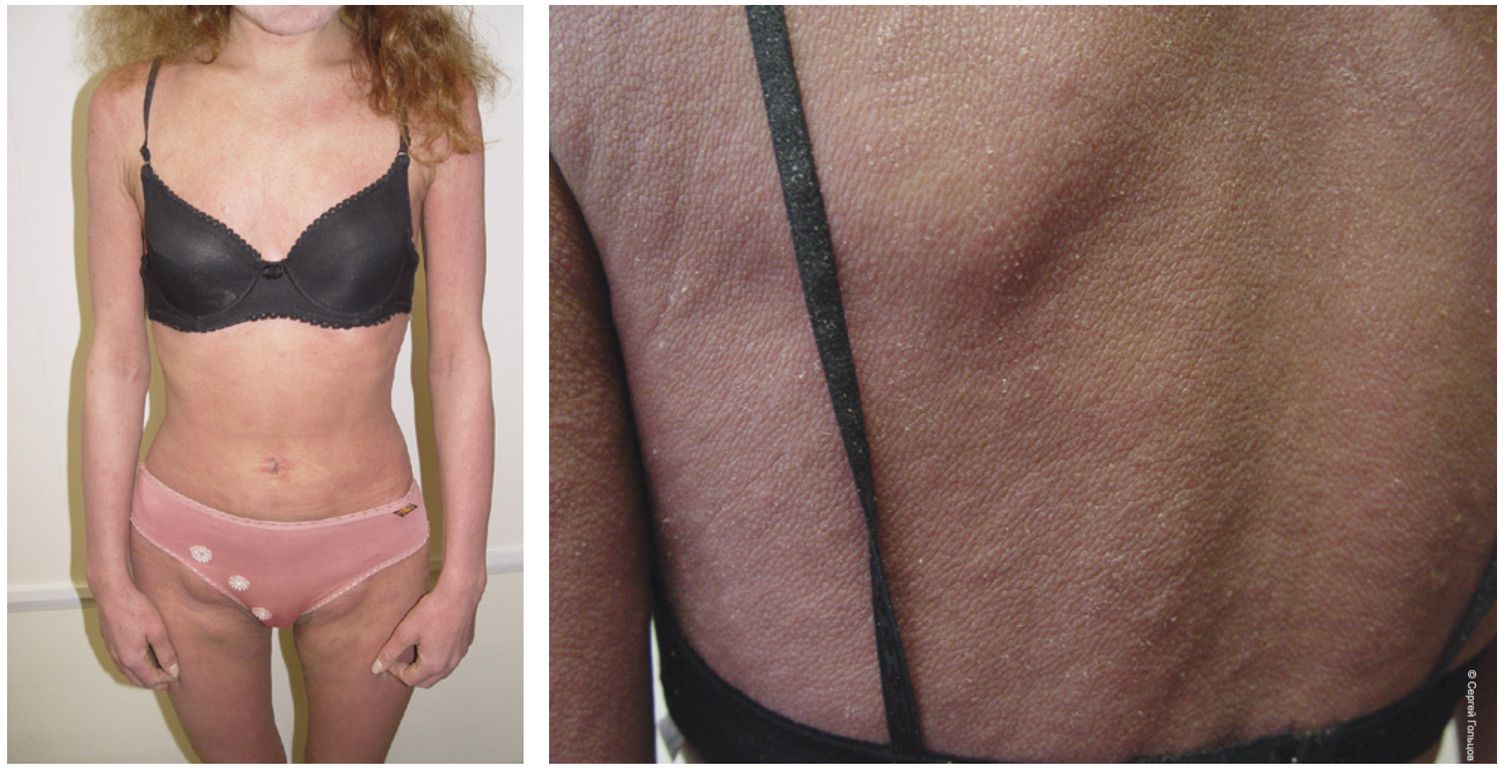
Figure 31. Clinical presentation of patient L., 26 years old, December 12, 2014, No. 11/2.
Based on the patient’s complaints, medical history, clinical findings, and examination results, the diagnosis of severe atopic dermatitis with erythroderma was established.
According to current dermatology clinical guidelines, histological examination of skin biopsy specimens may be performed in complex cases requiring differential diagnosis of atopic dermatitis. A skin biopsy was therefore obtained to assess the functional characteristics of epidermal and dermal cells within the lesion (Table 3). Because the pathological process involved virtually the entire skin surface, the biopsy was collected from the upper outer quadrant of the right buttock.
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Показатели, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
79,0 11,8 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
51,0 34,2 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
47,0 0 9,9 11,4 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
0,9 0 25,1 1,0 5,2 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
75,4 8,9 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
6,9 1,0 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
25,0 15,0 8,0 6,0 11,0 |
|
Жизнеспособность, % |
94 |
The obtained data demonstrated pronounced manifestations of systemic inflammation and immune dysregulation at the level of the skin cellular phenotype. Despite preservation of overall cellular viability (94%), a marked shift of the phenotypic balance toward activation of inflammatory and antigen-presenting cell populations was observed, accompanied by relative suppression of regenerative and regulatory compartments.
Keratinocytes (CD49f⁺ HLA-DR⁺).
Keratinocytes accounted for 79% of the analyzed cells, with 11.8% represented by activated HLA-DR–expressing forms. This finding indicates stress-induced epidermal activation and impairment of skin barrier function. Keratinocytes appear to acquire immunocompetent properties, participating in antigen presentation and maintenance of inflammation.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺).
Fibroblasts constituted 51% of the cellular population, of which 34.2% displayed an activated phenotype (CD44⁺ CD80⁺). This finding reflects marked activation of the dermal matrix, suggesting an attempt by the tissue to compensate for chronic inflammation through enhanced extracellular matrix synthesis and remodeling.
Langerhans Cells (CD207⁺).
The proportion of Langerhans cells was 47%, with approximately 11% expressing activated CD80⁺ HLA-DR⁺ phenotypes. This pattern indicates high antigen-presenting activity and involvement in the Th2-mediated inflammatory cascade. The coexistence of multiple functional subtypes (activated and non-activated, HLA-DR positive and negative) suggests heterogeneous activation of cutaneous immune surveillance mechanisms.
Endothelial Cells (CD146⁺).
Although endothelial cells represented only approximately 0.9% of the population, the CD54⁺ HLA-DR⁻ and CD34⁺ subpopulations accounted for 25.1% and 5.2%, respectively, indicating substantial vascular participation in the inflammatory process. Endothelial activation and the development of a microvascular inflammatory component are characteristic features of erythroderma.

Mast Cells (CD249⁺ CD63⁺).
The total mast-cell population reached 75.4%, of which 8.9% were activated. This finding reflects pronounced mast-cell degranulation with release of inflammatory mediators, including histamine and tryptase, correlating clinically with pruritus, erythema, and vascular hyperreactivity.
Monocytes (CD45⁺ CD14⁺ HLA-DR⁺).
Monocytes represented 6.9% of the population, with activated forms accounting for 1.0%. This pattern suggests moderate infiltration by monocyte/macrophage-lineage cells and is consistent with chronic, relatively stable inflammation rather than acute inflammatory escalation.
Epidermal Lymphocytes.
The cutaneous immune profile was Th2-dominant, accompanied by a moderate increase in NK cells, indicating recruitment of innate immune mechanisms. The Th/CD8 ratio remained approximately 1.9, demonstrating relative preservation of T-cell balance but with a shift toward helper-cell predominance characteristic of atopic disease.
The obtained cytoimmunogram corresponds to a highly active inflammatory skin phenotype characterized by:
Such a phenotype is associated with chronic atopic dermatitis during an acute exacerbation complicated by erythroderma.
For evaluation of therapeutic efficacy, the practical significance of this observation lies in establishing the baseline phenotype of inflammatory infiltrate cells within the skin before treatment. This baseline subsequently served as a reference point for monitoring phenotypic changes during inpatient therapy, which consisted of intravenous methylprednisolone (250 mg in 200 mL saline, five infusions), 30% sodium thiosulfate (10 mL intravenously daily, 15 administrations), 10% calcium gluconate (10 mL intramuscularly daily, 15 administrations), promethazine (25 mg once daily for 5 days), followed by clemastine (1 mg three times daily for 10 days), together with topical mometasone furoate, emollients, and irrigation of excoriated lesions with 0.05% chlorhexidine bigluconate solution. The clinical response was substantial and manifested as a marked reduction in pruritus and cutaneous infiltration.
Analysis of mast-cell subpopulations before treatment and 1.5 months after initiation of therapy revealed a pronounced decrease in activated mast cells (CD249⁺ CD63⁺ phenotype) within the inflammatory focus (Figure 32).
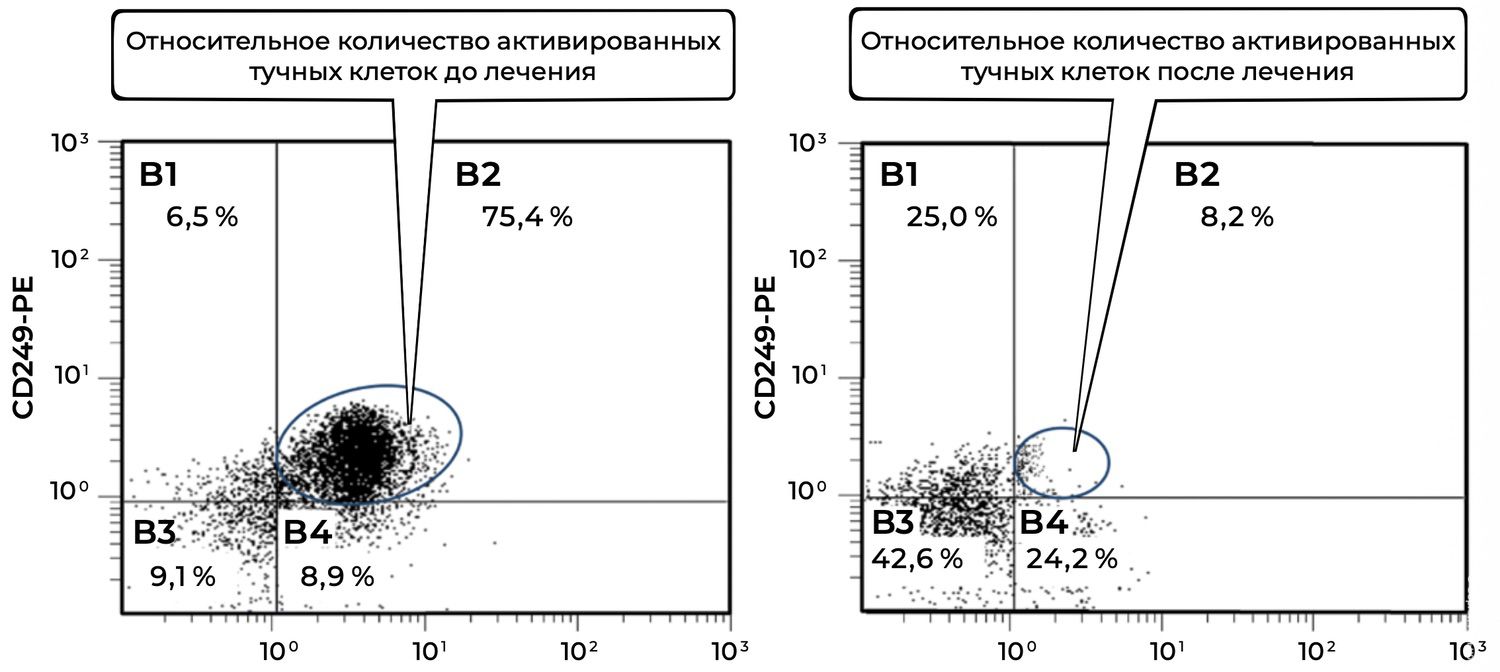
Figure 32. Comparative analysis of the relative proportion of viable activated mast cells (CD249⁺ CD63⁺ phenotype) in skin-biopsy cell suspensions obtained from patient L., 26 years old, on December 12, 2014 (No. 11/2), and January 25, 2015 (No. 11/5).
The comparative evaluation of viable activated mast cells (CD249⁺ CD63⁺) before and after treatment demonstrated the following.
The analyzed population consisted of viable activated mast cells identified by expression of two markers:
CD249 (p75/NGFR) — a surface marker of viable mast cells;
CD63 — an activation marker associated with degranulation and the release of inflammatory mediators such as histamine and tryptase.
Mast-cell activation decreased by a factor of 9.2, from 75.4% to 8.2%. Such a marked reduction indicates the effectiveness of the therapeutic regimen in stabilizing mast-cell activity, suppressing degranulation, and attenuating the inflammatory component of the disease.
Additional observations from the cytometric plots included:
B3 quadrant (CD249⁻ CD63⁻) — non-specific or unidentified cells:
Before treatment: 9.1%
After treatment: 42.6%
This nearly fivefold increase suggests that a proportion of previously activated cells may have transitioned into a non-activated state.
B4 quadrant (CD249⁺ CD63⁻) — viable but non-activated mast cells:
Before treatment: 8.9%
After treatment: 24.2%
This 2.7-fold increase confirms that treatment preserved mast-cell viability while effectively suppressing cellular activation.
Thus, at baseline the patient exhibited pronounced inflammatory activation of cutaneous mast cells. Following therapy, the proportion of CD63⁺ activated mast cells declined dramatically, while the proportion of viable non-activated mast cells increased. These findings demonstrate not only clinical improvement but also objective immunocytological evidence of therapeutic efficacy.
The application of flow cytometry in practical dermatology substantially enhances diagnostic precision by enabling multiparametric analysis of skin-cell populations from a single patient sample. Equally important, it allows dynamic monitoring of phenotypic changes over time. This was illustrated by serial assessment of fibroblast activation before treatment, during therapy, and after clinical improvement, demonstrating a progressive decline in activated fibroblasts within the inflammatory infiltrate (Figure 33).
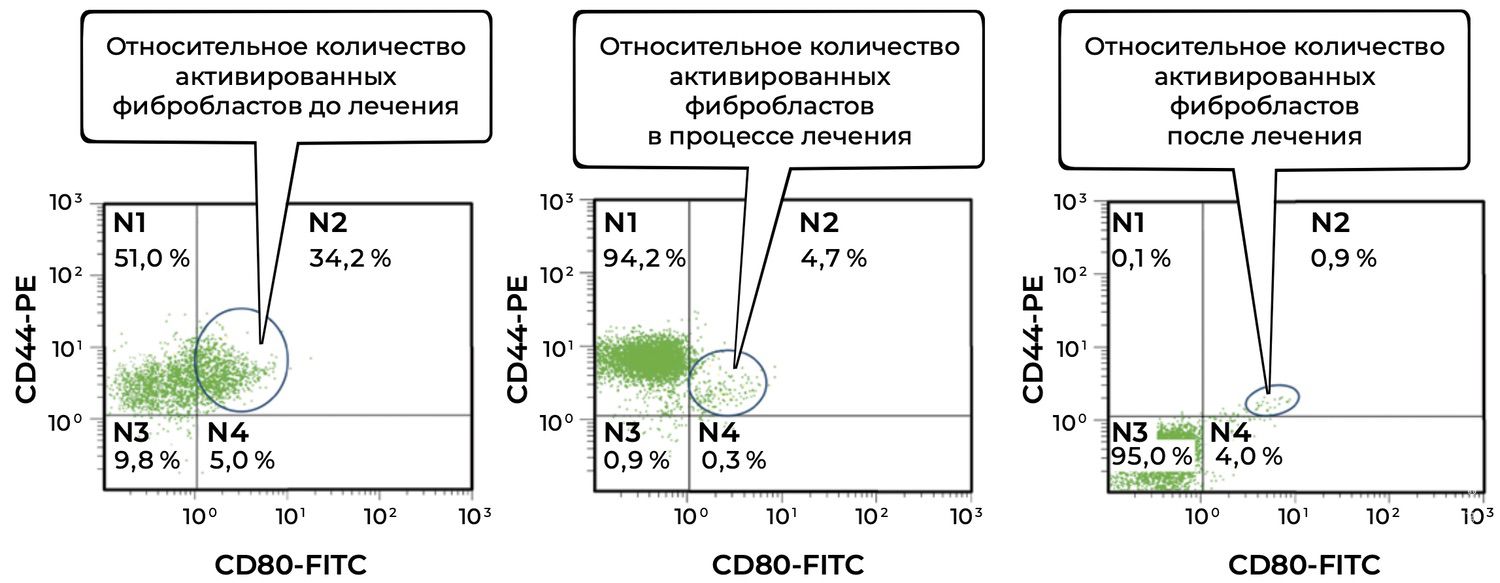
Figure 33. Comparative analysis of the relative proportion of viable activated fibroblasts in skin-biopsy cell suspensions from patient L., 26 years old, obtained on December 12, 2014 (No. 11/2), January 10, 2015 (No. 11/4), and January 25, 2015 (No. 11/5).
The dynamics of viable activated fibroblasts (CD45⁻ CD14⁻ CD44⁺ CD80⁺ phenotype) throughout treatment were as follows.
A 7.3-fold reduction was observed during treatment (34.2% → 4.7%).
A 38-fold reduction was observed by the end of therapy (34.2% → 0.9%).
This pronounced decline indicates strong anti-inflammatory and reparative effects of treatment.
Additional observations included:
N1 population (CD44⁺ CD80⁻):
Before treatment: 51.0%
During treatment: 94.2%
After treatment: 0.1%
This pattern suggests a temporary expansion of viable but non-activated fibroblasts during the reparative phase, followed by normalization of the cellular composition.
N3 population (CD44⁻ CD80⁻):
After treatment: 95.0%
Predominance of the N3 population at the final assessment reflects restoration of a near-normal cellular profile.
Thus, the patient initially demonstrated marked fibroblast activation, likely secondary to chronic inflammatory injury. Treatment resulted in near-complete elimination of the activated CD44⁺CD80⁺ phenotype, indicating suppression of the fibrotic cascade, stabilization of extracellular matrix remodeling, and completion of the reparative phase.
The status of keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) within inflammatory infiltrates before treatment, during therapy, and after treatment is shown in Figure 34.
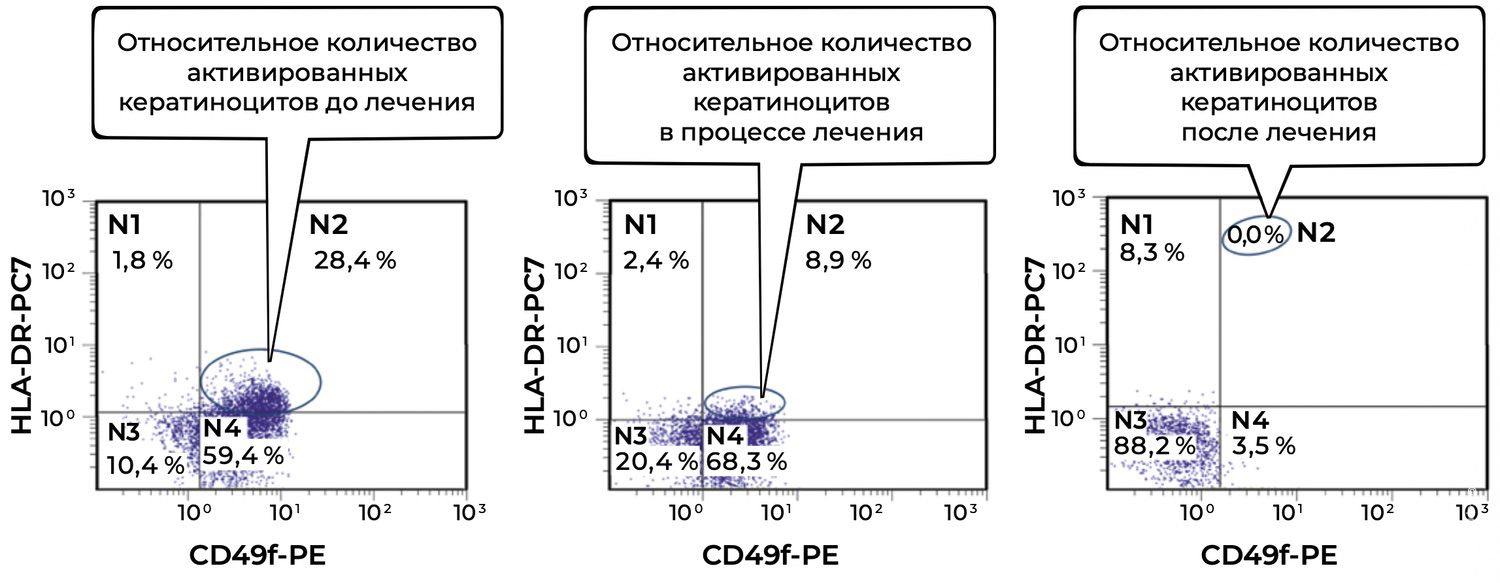
Figure 34. Comparative analysis of the relative proportion of viable activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in skin-biopsy cell suspensions from patient L., 26 years old, obtained on December 12, 2014 (No. 11/2), January 10, 2015 (No. 11/4), and January 25, 2015 (No. 11/5).
Before treatment, a high proportion of activated keratinocytes (28.4%) was detected, indicating pronounced epidermal activation and immune stress within the skin.
During treatment, activation decreased substantially to 8.9%, reflecting a favorable tissue response and attenuation of inflammatory activity.
After treatment, activated keratinocytes were no longer detected (0.0%), suggesting restoration of physiological epidermal homeostasis, normalization of immune balance, and resolution of active inflammation.
Thus, the reduction in activated keratinocytes (CD49f⁺ HLA-DR⁺) from 28.4% to 0.0% during treatment demonstrates a clear anti-inflammatory effect and restoration of the normal phenotypic status of epidermal cells. These findings support the use of this parameter as a reliable biomarker of keratinocyte inflammatory activity.
The above phenotypic findings corresponded closely with the positive clinical dynamics of the SCORAD index, which decreased from 82 at baseline to 41 one month after initiation of therapy and to 29 three weeks later. These results provide objective evidence of therapeutic effectiveness. The patient was subsequently discharged for outpatient follow-up treatment.
This case demonstrates the utility of skin-cell phenotypic analysis for the precision diagnosis of atopic dermatitis in adults. It also illustrates the value of skin cytoimmunograms as objective reference points for assessing the status of inflammatory skin infiltrates before treatment, during therapy, and after achievement of clinical remission, thereby complementing conventional visual and subjective measures of treatment response.
Patient M., a 57-year-old woman, presented to the Tyumen Regional Dermatovenerologic Dispensary with complaints of blisters appearing on the skin of the trunk and extremities, accompanied by mild pruritus in the evening. She considered herself ill for approximately six months, beginning when, without any apparent cause, she noticed the formation of small blisters in the oral cavity. At that time, she did not attach significance to these lesions.
One month later, against a background of worsening general well-being, she developed pruritus, erythematous macules, and small vesicles with turbid contents on the trunk and in the axillary regions. She self-treated with “various ointments from the home medicine cabinet,” but new lesions appeared on the chest and back and rapidly increased in size. Her past medical, hereditary, and allergic history was unremarkable. She denied recent antibiotic use.
At presentation to the outpatient clinic, the following differential clinical diagnosis was formulated: pemphigus vulgaris? pemphigus foliaceus? dermatitis herpetiformis Duhring?
Due to the widespread distribution and severity of the pathological process, the patient was hospitalized in the inpatient department of the Tyumen Regional Dermatovenerologic Dispensary. Examination of Tzanck smears for acantholytic cells and diagnostic biopsy of the skin of the back, taken from the marginal zone of an erosive surface, were prescribed.
Upon admission, the patient’s condition was classified as severe. She had a hypersthenic body constitution and grade III obesity. On auscultation, vesicular breathing was noted with moist rales. Respiratory rate was 20 breaths per minute. Heart sounds were clear and rhythmic; heart rate was 88 beats per minute, and blood pressure was 140/90 mmHg. The tongue was dry, with white coating along the lateral borders. The abdomen was soft and non-tender on palpation. The liver was enlarged by 2 cm below the costal margin; the spleen was not enlarged. Bowel movements were irregular, with a tendency toward constipation. Costovertebral angle tenderness was negative bilaterally. Urination was painless. The thyroid gland was not visually enlarged and was non-tender on palpation.
At admission, the pathological process was widespread, involving the skin of the trunk and both upper and lower extremities. It was represented by multiple round erythematous-squamous lesions of pink and red color, with rare brown hyperpigmented macules. On the surface of the lesions, predominantly on the back and chest, grouped vesicles with serous contents were observed on a hyperemic background; these measured up to 1 cm in diameter, had a tense and firm roof, and contained serous fluid. On the trunk, lateral surfaces, buttocks, lumbar region, and extremities, confluent erosions forming extensive foci were noted, with crusts and scales on their surface. Nikolsky’s sign was positive. Skin appendages were not involved. In the oral cavity, erosions and ulcers with white coating were present and painful upon pressure with a spatula.
Comprehensive clinical and laboratory evaluation revealed a positive Tzanck smear for acantholytic cells. Peripheral blood showed leukocytosis of 14.4 × 10⁹/L, lymphopenia of 1.19 × 10³/µL, and alanine aminotransferase level of 69 U/L. Direct immunofluorescence with antibodies to IgG, IgA, and IgM performed on a biopsy specimen of apparently unaffected skin revealed pronounced IgG deposition in the intercellular spaces throughout all layers of the epidermis. No IgA or IgM deposition was detected in skin structures.
These findings formed the basis for the diagnosis: pemphigus vulgaris, severe course. PDAI score: 190 points (Figure 35).
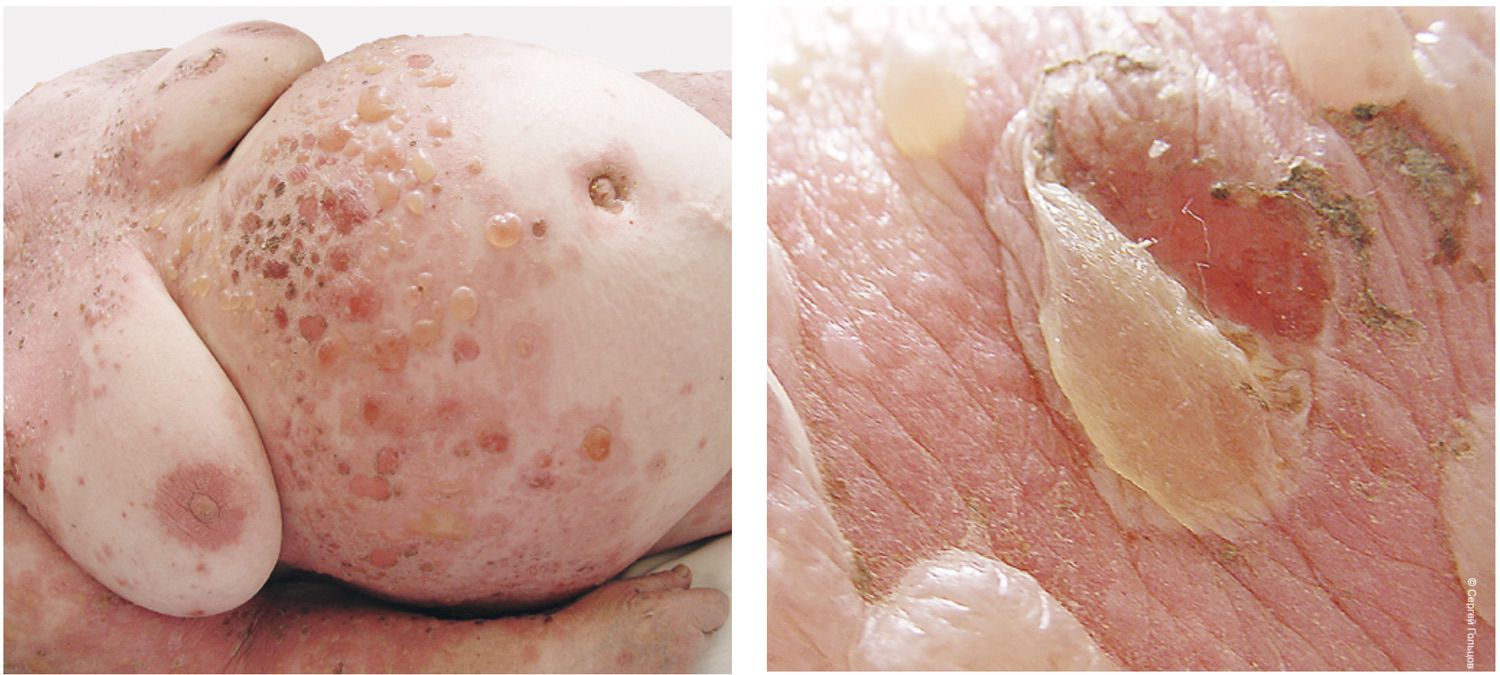
Figure 35. Clinical presentation of the local status of patient M., 57 years old, November 12, 2016.
During inpatient treatment, the patient received prednisolone 100 mg daily, calculated at 1 mg/kg body weight, throughout the 15-day hospitalization period, followed by an immediate one-third dose reduction and subsequent tapering by 5 mg per week. Additional therapy included essential phospholipids, 5 mL intravenously by bolus injection, No. 10; Panangin, one tablet three times daily; calcium D3, one tablet three times daily; omeprazole 20 mg, one capsule twice daily; topical therapy with mometasone furoate ointment; and irrigation with methylene blue solution.
The skin cytoimmunogram of the marginal zone of erosions is presented below (Table 4).
Table 4. Skin Cytoimmunogram of Patient M., 57 Years Old, November 12, 2016, No. 138/4
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Показатели, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
41,2 29,7 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
54 0,6 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
57 23,9 7,0 0,5 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
0,9 4,2 12,5 0 0,2 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
2,6 1,9 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
4,0 2,5 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
15 11 2 5 14 |
|
Жизнеспособность, % |
92 |
The cellular suspension obtained from the biopsy specimen of the marginal zone of erosion demonstrated cell viability of 92%, which ensured the reliability of phenotyping. The cytoimmunogram profile reflected an autoimmune inflammatory phenotype characterized by a combination of epidermal hyperactivation, high antigen-presenting activity, a vascular inflammatory component, and moderate participation of innate immunity. This phenotype corresponded to the active phase of an autoimmune process with focal regenerative compensation.
The analysis of cellular subpopulations is presented below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
Keratinocytes accounted for 41.2% of the population, with a high activation level of 29.7%. These data indicate profound epidermal activation associated with autoimmune attack against desmosomal proteins, particularly desmogleins. HLA-DR expression on keratinocytes indicates their involvement in antigen-presenting function, a key pathogenetic mechanism of pemphigus that contributes to maintenance of the local autoimmune response.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺).
Fibroblasts constituted 54% of the population, while activated forms accounted for only 0.6%. This low level of activation indicates suppression of reparative mechanisms within the lesion, likely due to continuous inflammatory pressure and the toxic effect of autoantibodies. At the same time, preservation of the overall fibroblast fraction above baseline reflects tissue readiness for regeneration once the inflammatory stimulus is removed.
Langerhans Cells (CD207⁺).
The total proportion of Langerhans cells was 57%, with activated forms (CD80⁺ HLA-DR⁺) reaching 23.9%, indicating marked hyperactivation of the antigen-presenting compartment. A multipolar distribution of phenotypes (HLA-DR⁺/HLA-DR⁻) was observed, corresponding to heterogeneity of the immune response within the marginal zone of erosions: a combination of actively antigen-presenting cells and cells in a state of functional exhaustion. This pattern is typical of autoimmune processes characterized by cyclical activity.
Endothelial Cells (CD146⁺ CD54⁺ CD34⁺).
The cumulative proportion was 17.8%, with CD54⁺ HLA-DR⁻ cells accounting for 12.5%. This profile reflects increased vascular permeability and endothelial activation, clinically manifesting as exudation and blister formation. Minimal presence of CD34⁺ cells (0.2%) indicates limited angiogenesis, which may explain delayed epithelialization of erosions.
Mast Cells (CD249⁺ CD63⁺).
Mast cells accounted for 2.6% of the population, with activated forms representing 1.9%. Moderate mast-cell degranulation was observed, supporting inflammation through the release of histamine, tryptase, and TNF-α. This ratio is typical of a chronic, non-acute course of pemphigus.
Monocytes (CD45⁺ CD14⁺ HLA-DR⁺).
The total monocyte fraction was 4%, with activated forms accounting for 2.5%. This reflects local macrophage-lineage infiltration and activation of the phagocytic compartment responsible for clearance of autoantibodies and cellular debris. The presence of HLA-DR⁺ monocytes underscores the participation of this compartment in antigen presentation and maintenance of the autoimmune reaction.
Epidermal Lymphocytes:
Total T cells (CD45⁺ CD3⁺): 15%;
T-helper cells (CD4⁺): 11%;
Cytotoxic T cells (CD8⁺): 2%;
B cells (CD19⁺): 5%;
NK cells (CD16⁺ CD56⁺): 14%.
The immune profile was characterized by predominance of the Th compartment (CD4⁺), with low levels of cytotoxic T cells (CD8⁺), which is typical of autoantibody-dependent inflammation. The high proportion of NK cells (14%) reflects activation of innate immunity in response to epidermal destruction and blister formation.
The phenotypic pattern indicates an autoimmune-inflammatory type of injury with epidermal hyperactivation, pronounced antigen presentation, and involvement of vascular endothelium. At the same time, a moderate reparative tendency was observed, reflected by activation of CD14⁺ HLA-DR⁺ cells and the presence of CD34⁺ cells. This allows the phenotype to be classified as a transitional inflammatory–regenerative type.
The skin cytoimmunogram of patient M. reflects an active autoimmune inflammatory phenotype typical of pemphigus vulgaris:
enhanced activation of keratinocytes and Langerhans cells;
an antigen-presenting cascade involving HLA-DR⁺ cells;
endothelial reactivity (CD54⁺ CD146⁺);
participation of mast cells and macrophages in sustaining inflammation;
a Th-dominant profile with a high proportion of NK cells.
The profile corresponds to the active phase of pemphigus within the erosive zone. With therapy aimed at reducing antigen presentation and restoring intercellular adhesion, a transition toward a remission-type skin cytoimmunogram may be expected.
In the presented example, the quantitative and functional status of the principal skin-cell subpopulations is documented in a specific patient, with her unique characteristics of both the skin and the organism as a whole. In effect, the photograph of the rash provides information about local clinical status but does not contain information about the cells forming the observed morphological elements. Meanwhile, according to clinical recommendations, the choice and dosage of systemic glucocorticosteroids in pemphigus depend on the severity and localization of clinical manifestations, the form and duration of the disease, and data on the effectiveness of previous therapy.
Thus, this choice is determined by the dermatologist’s subjective perception and clinical experience rather than by an objective quantitative and functional characterization of the patient’s skin cells. This is evidently related to the fact that both topical and systemic therapies remain universal and do not account for the actual cellular state of the skin in an individual patient. Yet such differences do exist.
For example, using only one parameter of the skin cytoimmunogram, it is possible to trace that the proportion of keratinocytes (CD49f⁺) in the biopsy specimen from the patient’s skin infiltrate was 41.2%, of which 29.7% were activated (CD49f⁺ HLA-DR⁺). After treatment according to the Federal Clinical Guidelines for Dermatology (2016), these values were 59.6% and 37.0%, respectively (Figure 36).
Figure 36. Comparative analysis of the relative proportion of viable activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient M., 57 years old, obtained on November 12, 2016 (No. 138/4), and November 27, 2016 (No. 138/5).
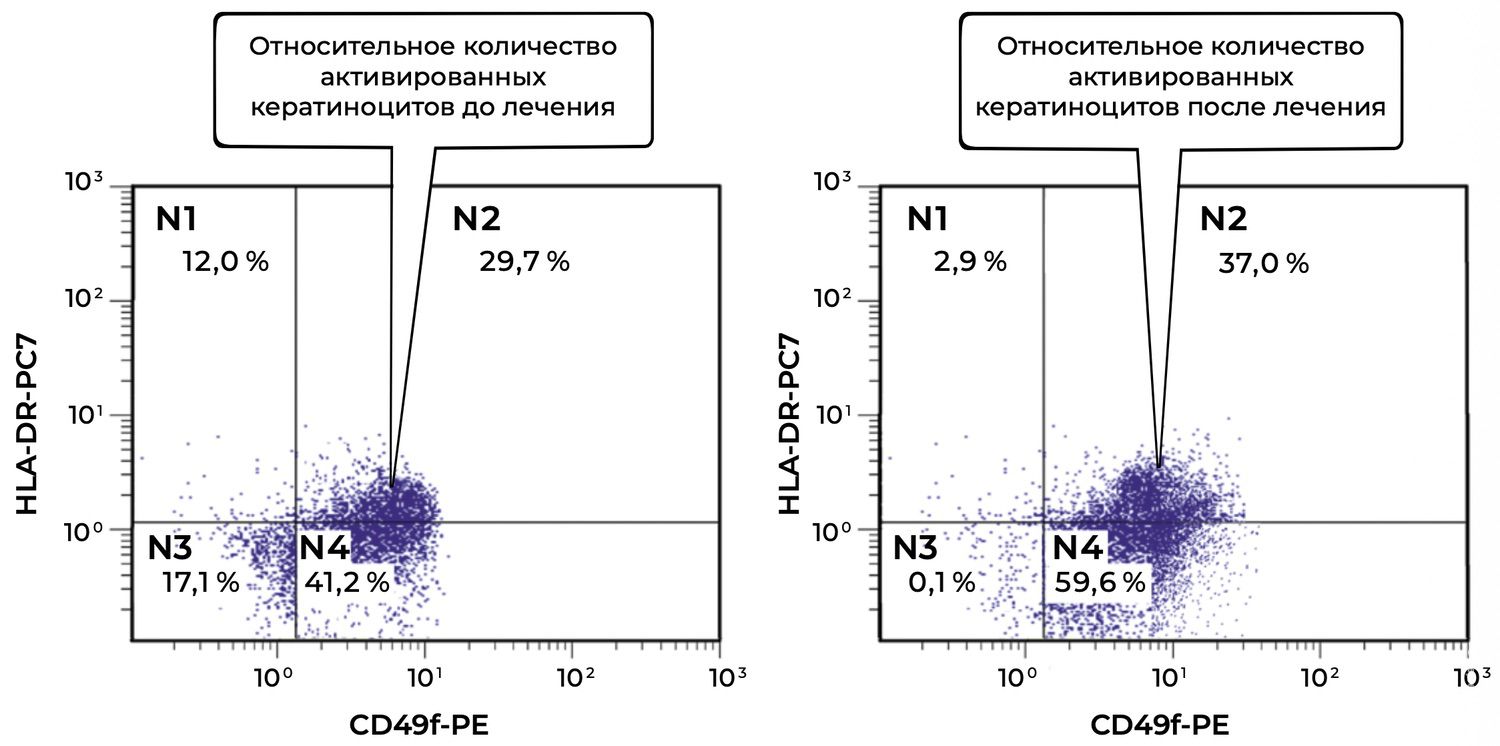
The comparative description of viable activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient M., based on the diagrams obtained before and after treatment, is as follows.
Before treatment, a high level of keratinocyte activation was observed: nearly one-third of epidermal cells (29.7%) expressed HLA-DR in combination with CD49f, corresponding to an inflammatory or immune-reactive state of the skin.
After treatment, contrary to expectation, the proportion of activated keratinocytes increased to 37.0%. This may indicate one of several possibilities:
enhancement of the inflammatory response during treatment, for example in the case of individual intolerance or reaction to treatment components;
a late reparative phase in which HLA-DR expression functions as a marker of regeneration rather than destruction;
insufficient efficacy of the therapy with respect to the inflammatory component, requiring reassessment of the treatment strategy.
Additional observations include the following.
The proportion of keratinocytes with a viable non-activated phenotype (CD49f⁺ HLA-DR⁻, region No. 4) also increased, from 41.2% to 59.6%, which may indicate compensatory proliferation of non-pathogenic epithelial cells.
Region No. 3, corresponding to the double-negative population, almost disappeared after treatment (17.1% → 0.1%). This may be interpreted as a reduction in the number of damaged and metabolically “silent” cells, which is generally a favorable sign.
Unlike in the previous case, the relative proportion of activated keratinocytes in this patient did not decrease but increased during treatment (29.7% → 37.0%). This requires comprehensive clinical interpretation: increased HLA-DR expression may be negative, as a sign of persistent inflammation, or neutral/positive, reflecting active regeneration. In this case, correlation with clinical findings, inflammatory cytokine levels, morphology, and overall therapeutic response may be recommended.
The proposed method of evaluating skin cytoimmunograms is not only a solution to an important problem in scientific research, but also allows the dermatologist to obtain objective information about the presence of an autoimmune process in the patient’s skin. These results may be used as an additional diagnostic criterion and as a means of improving therapeutic monitoring. As in most other skin diseases, histological examination of skin biopsy specimens is often performed for differential diagnosis. However, it is not used for treatment selection, which is striking, since the skin cytoimmunogram makes it possible to add a functional characterization of skin cells to the morphological description—especially important for assessing the actual state of the skin.
At discharge from the hospital, the patient demonstrated positive clinical dynamics, with regression of most lesions and formation of residual hyperpigmentation. On the skin of the trunk and upper and lower extremities, isolated firmly adherent dry crusts remained. The patient was discharged with clinical improvement for outpatient continuation of therapy. Recommendations included continuing prednisolone at a dose of 70 mg per day, with subsequent consideration of dose reduction based on clinical and laboratory data under the supervision of a dermatologist at the place of residence. Hepatoprotective agents, proton pump inhibitors, potassium and magnesium preparations, and a calcium–phosphorus metabolism regulator were also recommended.
Objectively, this was accompanied by a decrease in the number of activated intraepidermal macrophages (CD207⁺ HLA-DR⁺) in skin cytoimmunograms obtained on November 12, 2016 (initiation of treatment), and November 27, 2016 (at the time of the initial reduction of prednisolone dose from 100 mg/day to 70 mg/day) (Figure 37).
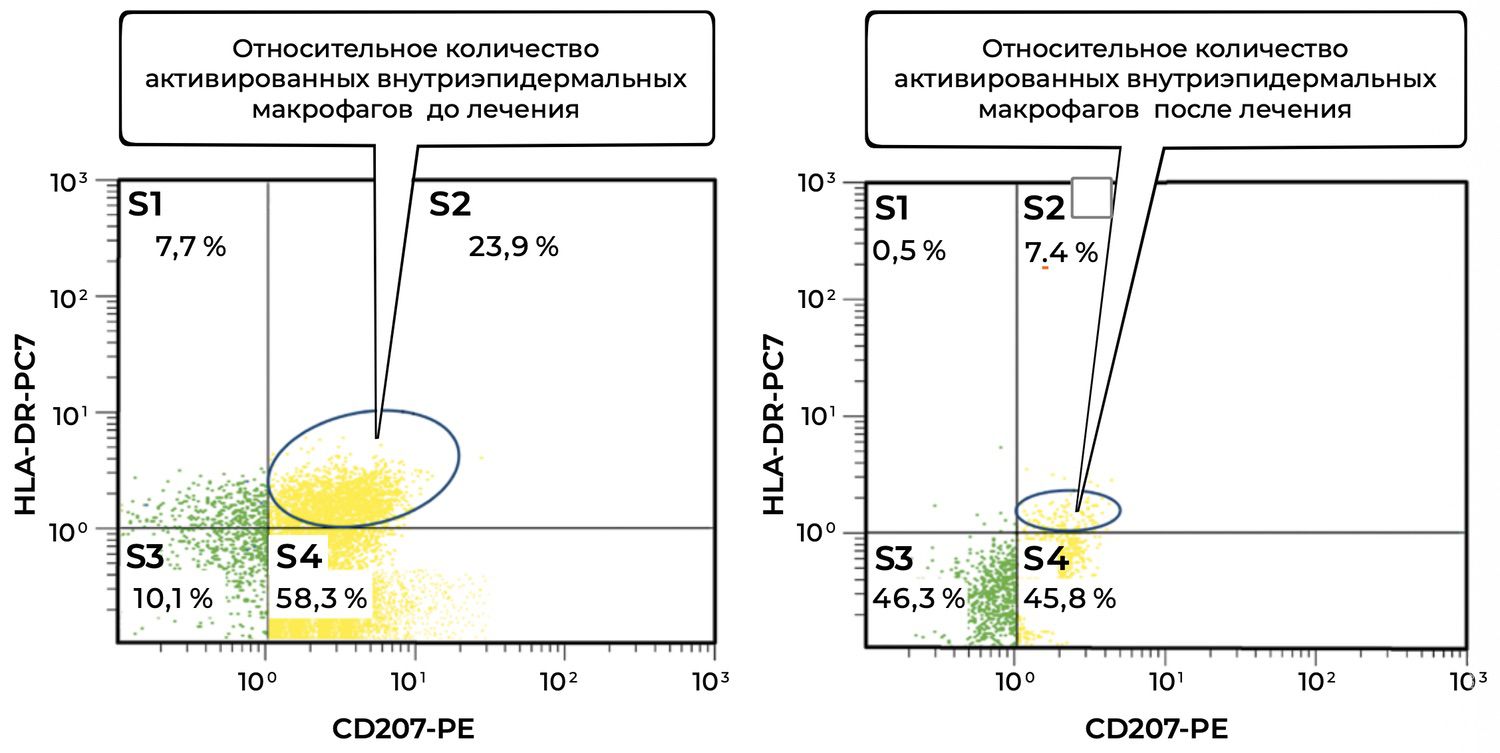
Figure 37. Comparative analysis of the relative proportion of activated intraepidermal macrophages (CD207⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient M., 57 years old, obtained on November 12, 2016 (No. 138/4), and November 27, 2016 (No. 138/5).
The comparative analysis of activated intraepidermal macrophages (CD207⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension before and after treatment is as follows.
Before treatment, the high proportion of activated intraepidermal macrophages (23.9%) indicated a pronounced inflammatory or immune response in the skin. CD207⁺ HLA-DR⁺ cells reflect functional activation of Langerhans antigen-presenting cells.
After treatment, a marked decrease in activated macrophages to 7.4% was observed. This may be interpreted as:
successful suppression of the inflammatory component by therapy;
and/or restoration of the physiological microenvironment of the epidermis;
possible return of macrophages to a “resting” or tolerant phenotype.
Additional zones provide further information.
The total proportion of CD207⁺ cells (S2 + S4) decreased from 82.2% (23.9% + 58.3%) to 53.2% (7.4% + 45.8%). Thus, the overall number of CD207⁺ macrophages also decreased, which may reflect not only deactivation but also reduced migration and presence of these cells within the lesion.
At the same time, the S3 population (CD207⁻ HLA-DR⁻) increased from 10.1% to 46.3%, which may indicate an increased presence of inactive or “reserve” cutaneous immune cells not involved in active inflammation.
Overall, the patient demonstrated a substantial reduction in activated intraepidermal macrophages CD207⁺ HLA-DR⁺ after treatment (23.9% → 7.4%). This may indicate the effectiveness of therapy aimed at modulating the cutaneous immune response and improving homeostasis of the epidermal immune microenvironment.
Two months after inpatient treatment, the daily dose of prednisolone was reduced from 70 mg to 30 mg. However, isolated erosions persisted, requiring stimulation of reparative processes. The skin cytoimmunogram showed a structural and functional deficit of reparative potential, determined by the level of endothelial cells (Figure 38).
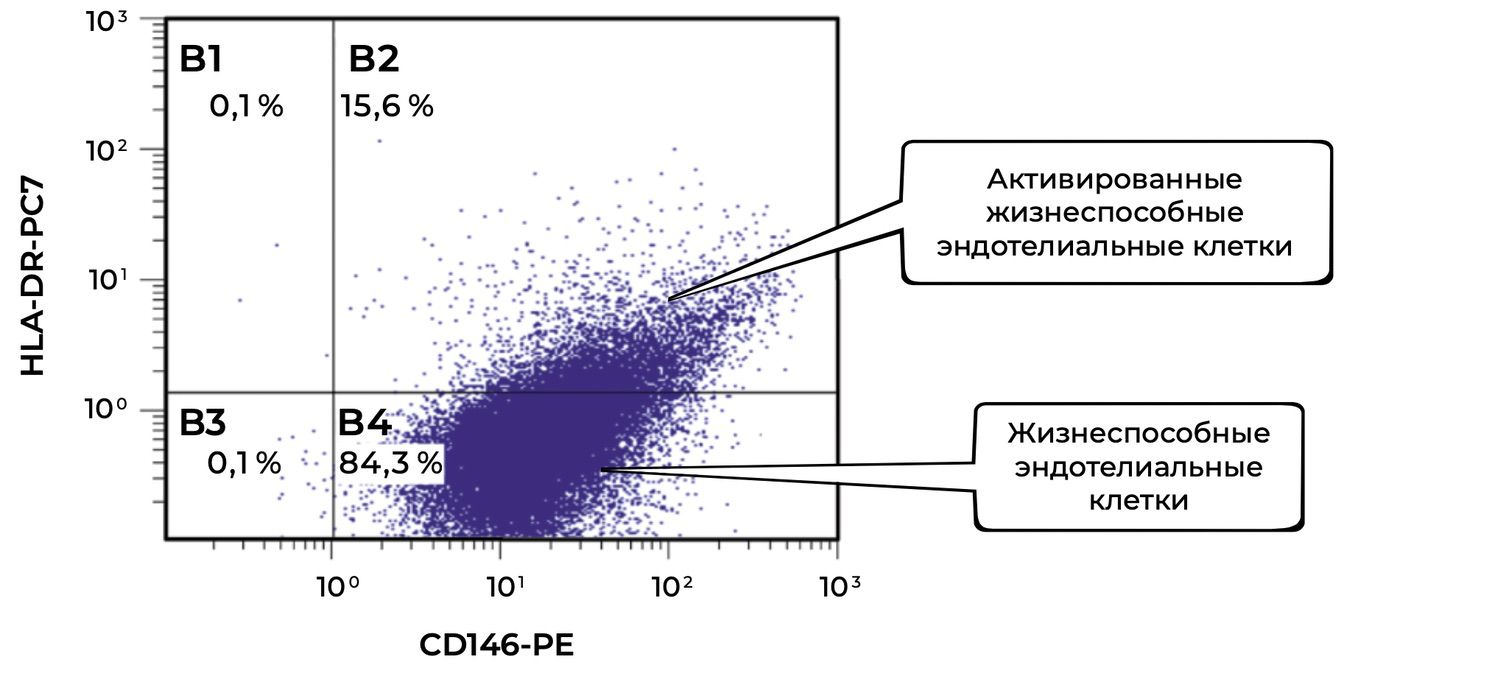
Figure 38. Comparative analysis of the relative proportion of viable endothelial cells (CD146⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient M., 57 years old, obtained on November 27, 2016 (No. 138/5).
The final description of viable endothelial cells (CD146⁺ phenotype) and their activated forms (CD146⁺ HLA-DR⁺ phenotype) in the patient after treatment, based on flow-cytometric analysis, is as follows.
The total proportion of viable endothelial cells (CD146⁺) in the suspension was 99.9%, indicating excellent preservation of the endothelial pool in the skin after treatment.
Of these, 15.6% were activated by HLA-DR expression, indicating:
preserved immunological competence of the endothelium;
moderate activation of antigen-presenting mechanisms;
participation in regulation of the local immune response and microvascular remodeling.
At the same time, the majority of cells (84.3%) remained viable but non-activated, which is characteristic of stable physiological endothelium not engaged in aggressive inflammation.
After treatment, the patient retained a high proportion of viable endothelial cells (99.9%), of which only 15.6% showed signs of activation. This indicates restoration of the microcirculatory bed, reduction of vascular activation, and normalization of the local cutaneous immune background. Taken together, these changes indicate positive dynamics and therapeutic effectiveness.
These circumstances may serve as a rationale for prescribing agents with a proven stimulatory effect on cell proliferation during the second and third phases of wound healing.
Clinical guidelines in dermatovenerology recommend that dermatologists diagnose lichen planus according to criteria observed during visual examination of the rash:
“Skin involvement in the typical form of lichen planus is characterized by flat papules measuring 2–5 mm in diameter, with polygonal outlines, central umbilication, a pinkish-red color with a characteristic violaceous or lilac hue, and a waxy sheen that is more evident under oblique illumination. Scaling is usually minimal, and scales are difficult to detach. On the surface of larger papules, especially after application of oil, a reticular pattern may be detected (Wickham’s striae). A characteristic feature of lichen planus is the tendency of lesions to appear in grouped arrangements, forming rings, garlands, and lines. Less commonly, papules coalesce to form plaques with a shagreen-like surface. New papules may develop around plaques, distributed with varying density. In most cases, the eruption is symmetrically localized on the flexural surfaces of the extremities, trunk, genitalia, and frequently on the oral mucosa. The palms, soles, and face are rarely involved. Subjectively, patients report pruritus. During exacerbations, the Koebner phenomenon is observed—the appearance of new papules at sites of skin trauma…”⁵⁹
The patient presented with a characteristic clinical picture (Figure 39).
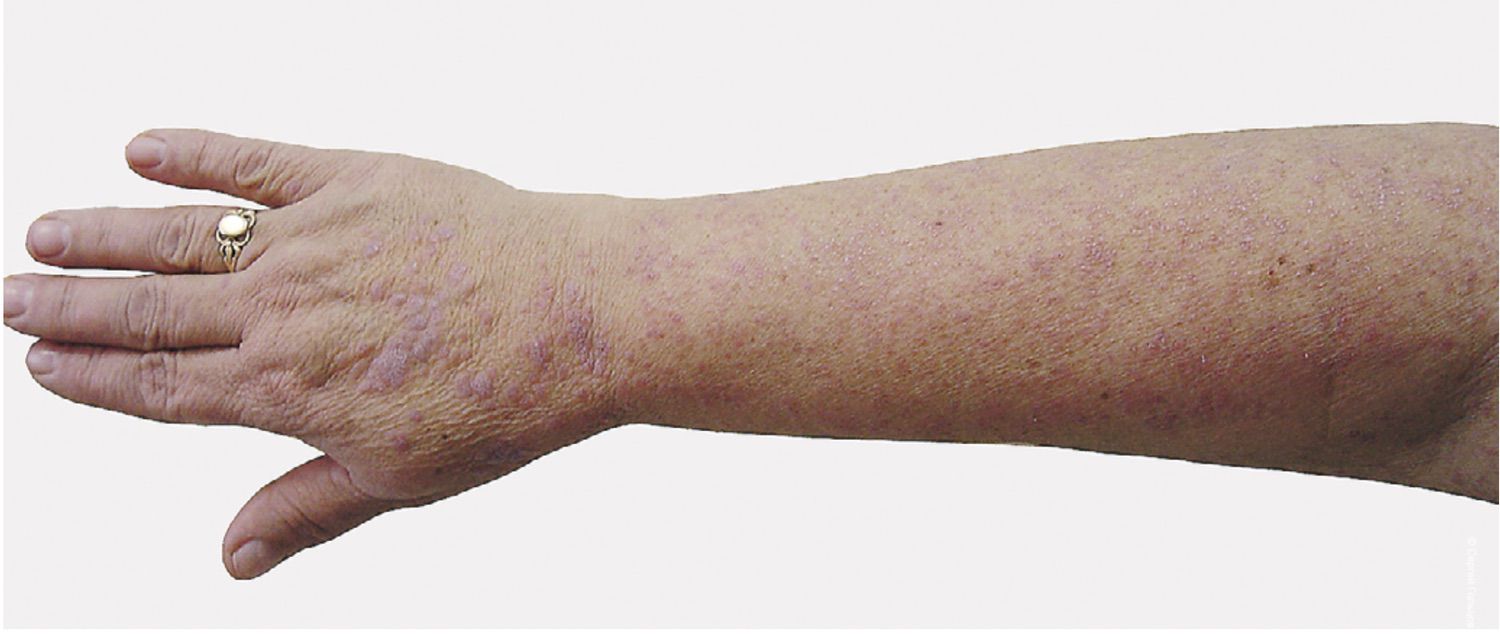
Figure 39. Clinical presentation of the local status of patient L., 52 years old, November 12, 2016.
At the same time, the cytoimmunogram of the patient’s skin infiltrates is presented (Table 5).
Table 5. Skin Cytoimmunogram of Patient L., 52 Years Old, November 12, 2016, No. 138/4
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Показатели, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
41 29,7 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
54 0,6 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
57 0,9 7,0 0,5 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
0,9 4,2 12,5 0 0,2 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
2,6 1,9 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
4,0 2,5 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
15 11 2 5 14 |
|
Жизнеспособность, % |
92 |
Cell viability was 92%, indicating good sample quality and absence of pronounced necrosis. The phenotypic profile reflects an immune-inflammatory type of cutaneous response, with active participation of antigen-presenting cells, the lymphocytic compartment, and a moderate vascular component. Unlike pemphigus, inflammation in this case is cell-mediated, without massive epidermolysis, but with pronounced cytotoxic and helper potential.
The analysis of cellular subpopulations is presented below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
The total proportion was 41%, with activated forms accounting for 29.7%. Increased HLA-DR expression indicates transformation of keratinocytes into “quasi–antigen-presenting” cells. This is one of the key pathogenetic mechanisms of lichen planus: keratinocytes become targets for CD8⁺ T cells, while simultaneously participating in maintenance of inflammation. The high proportion of activated cells confirms an active cytotoxic process within the epidermis.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺ / CD80⁺).
The total proportion was 54%, with activated forms accounting for 0.6%. This indicates a preserved but inactive dermal matrix. Fibroblasts are not involved in the acute inflammatory phase but provide structural support and participate in remodeling of the basement membrane, which is primarily affected in lichen planus.
Langerhans Cells (CD207⁺ CD80⁺ HLA-DR⁺).
The total proportion was 57%, while activated forms (CD80⁺ HLA-DR⁺) accounted for 0.9%, HLA-DR⁻ cells for 7%, and HLA-DR⁺ cells for 0.5%. The reduced level of HLA-DR⁺ cells indicates moderate antigen presentation in the context of a chronic course. Persistent stimulation of immune surveillance is observed, which explains the stability of lesions and the tendency toward recurrence.
Endothelial Cells (CD146⁺ / CD54⁺ / CD34⁺).
The cumulative proportion was approximately 18%, with predominance of CD54⁺ HLA-DR⁻ cells (12.5%). This is a sign of moderate endothelial activation accompanied by local vasodilation and lymphocytic infiltration into the dermoepidermal zone. The presence of CD34⁺ cells (0.2%) indicates weakly expressed angiogenesis.
Mast Cells (CD249⁺ / CD63⁺).
The total proportion was 2.6%, with activated forms accounting for 1.9%. Moderate mast-cell degranulation was observed, contributing to pruritus and maintenance of the local inflammatory response.
Monocytes (CD45⁺ CD14⁺ / HLA-DR⁺).
The total proportion was 4%, with activated forms accounting for 2.5%. This corresponds to moderate macrophage infiltration involved in destruction of damaged keratinocytes and clearance of the basement membrane zone.
Epidermal Lymphocytes:
Total T cells (CD45⁺ CD3⁺): 15%;
T-helper cells (CD4⁺): 11%;
Cytotoxic T cells (CD8⁺): 2%;
B lymphocytes (CD19⁺): 5%;
NK cells (CD16⁺ CD56⁺): 14%.
A Th-dominant profile is observed, with a low level of CD8⁺ cells, consistent with a chronic inflammatory pattern in which the cytotoxic compartment has already been engaged, while effector and memory T cells persist in the tissue. The increased NK-cell level (14%) indicates involvement of innate immune surveillance and correlates with persistence of lesions.
The phenotype corresponds to chronic cell-mediated inflammation with a moderate cytotoxic component, high keratinocyte activation, and moderate endothelial involvement. The epidermis remains hyperactive and immunocompetent, whereas dermal regenerative processes are suppressed. Overall, this profile may be classified as a Th1/Th17-dominant inflammatory type, typical of lichen planus.
The skin cytoimmunogram of patient L. demonstrates the following:
activated keratinocytes with HLA-DR expression involved in autoimmune inflammation;
persistent activity of Langerhans cells and the antigen-presenting compartment;
moderate activation of endothelial cells and macrophages;
a Th-dominant lymphocytic profile with NK-cell participation;
low fibroblast activation, reflecting chronicity and weak regeneration.
Thus, the phenotype corresponds to chronic Th1/Th17-mediated inflammation with cellular infiltration in the dermoepidermal zone, characteristic of lichen planus. The skin cytoimmunogram data confirm the need for immunomodulatory and anti-Th therapy to restore the structural and cellular equilibrium of the skin.
This example demonstrates the possibility of obtaining additional information about the cellular state of the observed inflammatory infiltrate both before and after treatment.
For instance, based on a single parameter of the skin cytoimmunogram, it is possible to trace that the proportion of activated keratinocytes in the biopsy specimen from the patient’s infiltrate was 29.7% before treatment and decreased to 8.9% after conventional therapy (Figure 40).
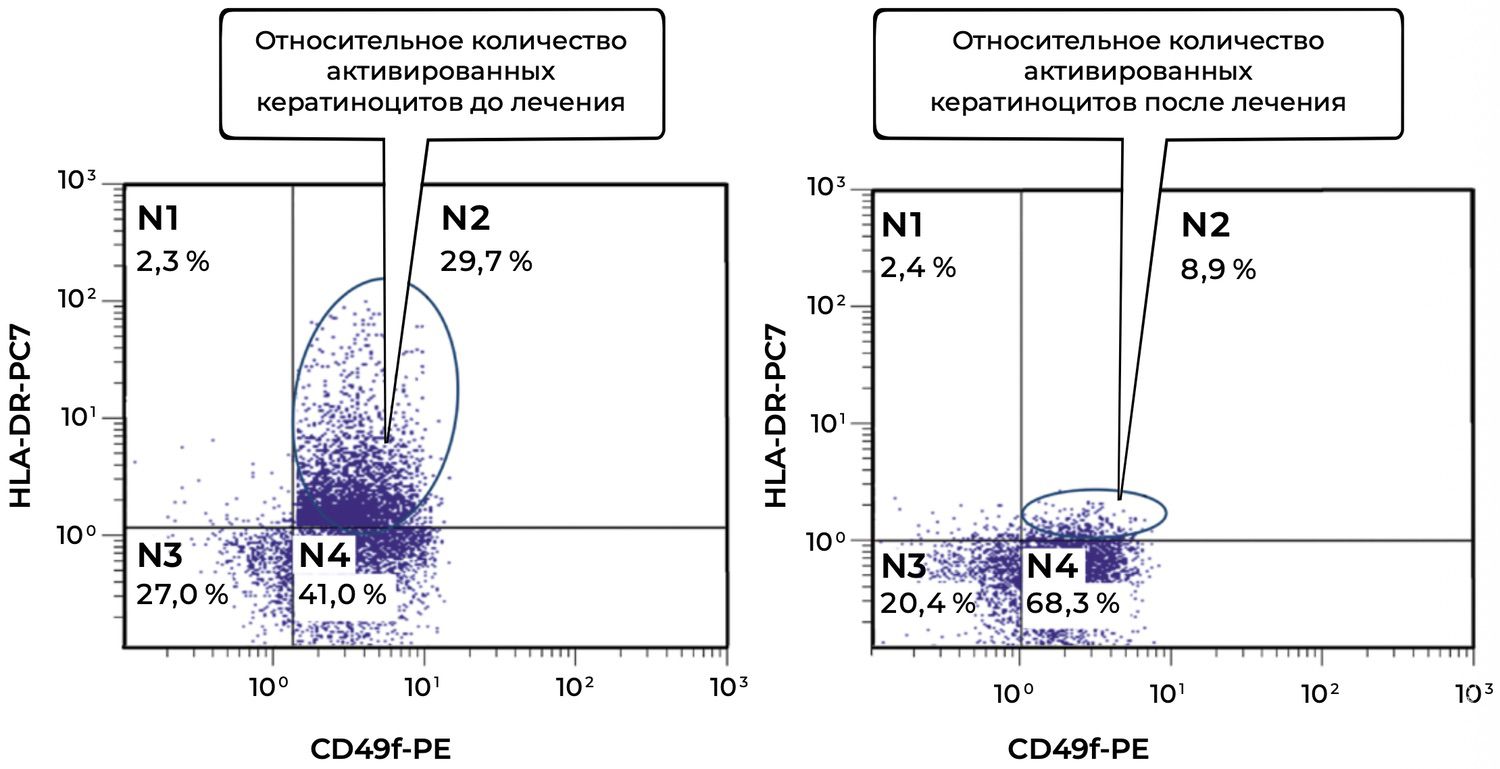
Figure 40. Comparative analysis of the relative proportion of viable activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient L., 52 years old, obtained on November 12, 2016 (No. 138/4), and November 27, 2016 (No. 138/5).
The cytoimmunogram presents the results of analysis of viable activated keratinocytes (CD49f⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of a patient with lichen planus before and after treatment.
Interpretation of Results
Before treatment, a substantial proportion of activated keratinocytes was observed (29.7%), which is typical of inflammatory dermatoses accompanied by hyperactivation of the epidermal compartment and acquisition of antigen-presenting function by keratinocytes.
After therapy, the proportion of activated keratinocytes decreased significantly to 8.9%, indicating reduced epidermal activation, suppression of the inflammatory process, and normalization of the functional status of the epidermis.
In parallel, the proportion of viable non-activated keratinocytes (N4) increased from 41.0% to 68.3%, indicating restoration of epidermal barrier structure and function, preservation of cellular viability, and regenerative potential of the skin.
Thus, during therapy, the patient demonstrated:
a pronounced reduction in activated keratinocytes (CD49f⁺ HLA-DR⁺);
an increase in viable non-activated keratinocytes, reflecting positive clinical and immunological dynamics.
This indicates the effectiveness of the treatment in reducing inflammation, restoring epidermal architecture, and suppressing the immune response at the skin level.
In this case, the method for obtaining skin cytoimmunograms allowed the dermatologist to collect additional objective information about the presence of an autoimmune proliferative process in the patient’s skin infiltrate and to take these findings into account during treatment.
According to the same clinical guidelines cited above, “psoriasis is a systemic immune-associated disease of multifactorial nature, with genetic factors playing a dominant role in its development. It is characterized by accelerated proliferation of epidermocytes and impaired differentiation, immune reactions in the dermis and synovial membranes, an imbalance between pro-inflammatory and anti-inflammatory cytokines and chemokines, and frequent pathological changes of the musculoskeletal system.” With respect to the immune status of the patient and the skin, “hereditary predisposition, dysfunction of the immune, endocrine, and nervous systems, adverse environmental influences, and frequent association with systemic diseases—including metabolic syndrome, type 2 diabetes mellitus, ischemic heart disease, arterial hypertension, hepatobiliary pathology, and others—are of clear significance.”
One such patient demonstrated the following skin cytoimmunogram parameters (Table 6).
Table 6. Skin Cytoimmunogram of Patient B., 51 Years Old, June 2, 2018, No. 89/2
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Показатели, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
65 54,4 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
20 2,9 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
58,3 23,9 14,4 9,8 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
1,4 0 19,9 0,4 1,9 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
1,6 0,3 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
6,6 0,1 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
15 10 5 7 8 |
|
Жизнеспособность, % |
89 |
Cell viability was 89%, indicating good biopsy quality and absence of massive necrosis. The immune-cellular skin profile demonstrated a highly active proliferative-inflammatory phenotype characteristic of the psoriatic process:
keratinocyte hyperactivation;
enhanced antigen presentation by Langerhans cells;
endothelial activation and lymphocytic infiltration of the Th17 type;
relatively low fibroblast and monocyte activity.
This phenotype corresponds to a stabilized but active type of psoriatic inflammation.
The analysis of cellular subpopulations is presented below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
The total proportion was 65%, with activated HLA-DR⁺ forms accounting for 54.4%, which substantially exceeds the expected baseline. This hyperactivation reflects an accelerated cell cycle and pathological epidermal proliferation typical of psoriasis. HLA-DR expression on keratinocytes indicates their involvement in antigen presentation, thereby sustaining chronic inflammation through a self-perpetuating cytokine loop involving the IL-23/Th17 axis.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺ / CD80⁺).
The total proportion was 20%, with activated forms accounting for 2.9%. Moderate activity of dermal fibroblasts was observed, reflecting their participation in matrix remodeling and maintenance of the inflammatory microenvironment. The reduced proportion of activated cells indicates an imbalance between inflammation and repair, typical of psoriatic hyperproliferative lesions.
Langerhans Cells (CD207⁺ CD80⁺ HLA-DR⁺).
The total proportion was 58.3%, while activated forms (CD80⁺ HLA-DR⁺/⁻) together exceeded 48%. This is a marked indicator of hyperactivity within the antigen-presenting compartment, consistent with the central role of Langerhans cells and dendritic cells in psoriasis pathogenesis, including initiation of the Th17 response and IL-23 production. The phenotype indicates persistent inflammation with active antigenic stimulation.
Endothelial Cells (CD146⁺ CD54⁺ CD34⁺).
The cumulative proportion was approximately 23%, with CD54⁺ HLA-DR⁻ cells accounting for 19.9%. This reflects pronounced endothelial activation, angiogenesis, and formation of the pathological microvascular network characteristic of psoriatic plaques. The presence of CD34⁺ cells (1.9%) indicates active angiogenesis. The endothelial component is one of the key morphological features of psoriasis.
Mast Cells (CD249⁺ CD63⁺).
The total proportion was 1.6%, with activated forms accounting for 0.3%. Mast-cell activity was low, which is typical of the chronic phase, in which inflammation is maintained primarily by lymphocytes and keratinocytes rather than mast-cell degranulation.
Monocytes (CD45⁺ CD14⁺ HLA-DR⁺).
The total proportion was 6.6%, with activated forms accounting for 0.1%. This reflects limited participation of the phagocytic compartment, characteristic of non-exudative inflammation. Macrophages appear to remain in a relatively resting state, consistent with the chronic nature of the process.
Epidermal Lymphocytes:
Total T cells (CD3⁺): 15%;
T-helper cells (CD4⁺): 10%;
Cytotoxic T cells (CD8⁺): 5%;
B lymphocytes (CD19⁺): 7%;
NK cells (CD16⁺ CD56⁺): 8%.
The immune profile was Th-dominant, with emphasis on the Th17/Th1 response and a moderate cytotoxic component (CD8⁺). The presence of both B cells and NK cells confirms involvement of a mixed adaptive–innate immune cascade.
The phenotype corresponds to chronic Th17-mediated inflammation with pronounced epidermal hyperactivation and an angiogenic component. Its principal features include:
hyperplasia and immune activation of keratinocytes (HLA-DR⁺);
strong antigen presentation (CD207⁺ CD80⁺ HLA-DR⁺);
active angiogenesis and endothelial response (CD54⁺ CD34⁺);
low fibroblast and monocyte activity;
a Th17-dominant cellular profile (CD4⁺ > CD8⁺, moderate NK-cell involvement).
This type of skin cytoimmunogram reflects a persistent inflammatory-proliferative skin phenotype maintained by the cytokine loop IL-23 → IL-17 → IL-22.
The skin cytoimmunogram of patient B. demonstrates:
massive activation of keratinocytes and disruption of their differentiation;
enhancement of Langerhans antigen-presenting cells;
a prominent vascular–angiogenic component;
Th17-mediated inflammation;
low activity of regenerative compartments.
The phenotype corresponds to the active stage of psoriasis, characterized by pronounced hyperproliferation and cytokine-dependent inflammation. Within the framework of phenotypic dermatology, this profile may be characterized as an inflammatory-proliferative type with an angiogenic dominance, determining the following therapeutic directions:
modulation of the Th17/IL-23 axis;
normalization of endothelial function;
stimulation of fibroblast-mediated regeneration.
It is customary to consider that “the diagnosis of psoriasis is established on the basis of the clinical picture of the disease, identification of the signs of the psoriatic triad, and the presence of the Koebner phenomenon,” and that the patient is concerned by “the appearance on the skin of pink-red papular elements with sharply defined borders, prone to coalescence and formation of plaques of various shapes and sizes, covered with silvery-white scales, predominantly on the scalp, extensor surfaces of the elbow and knee joints, lumbar and sacral regions, accompanied by pruritus of varying intensity” (Figure 41).
Figure 41. Clinical presentation of the local status of patient B., 51 years old, June 2, 2018, No. 89/2.
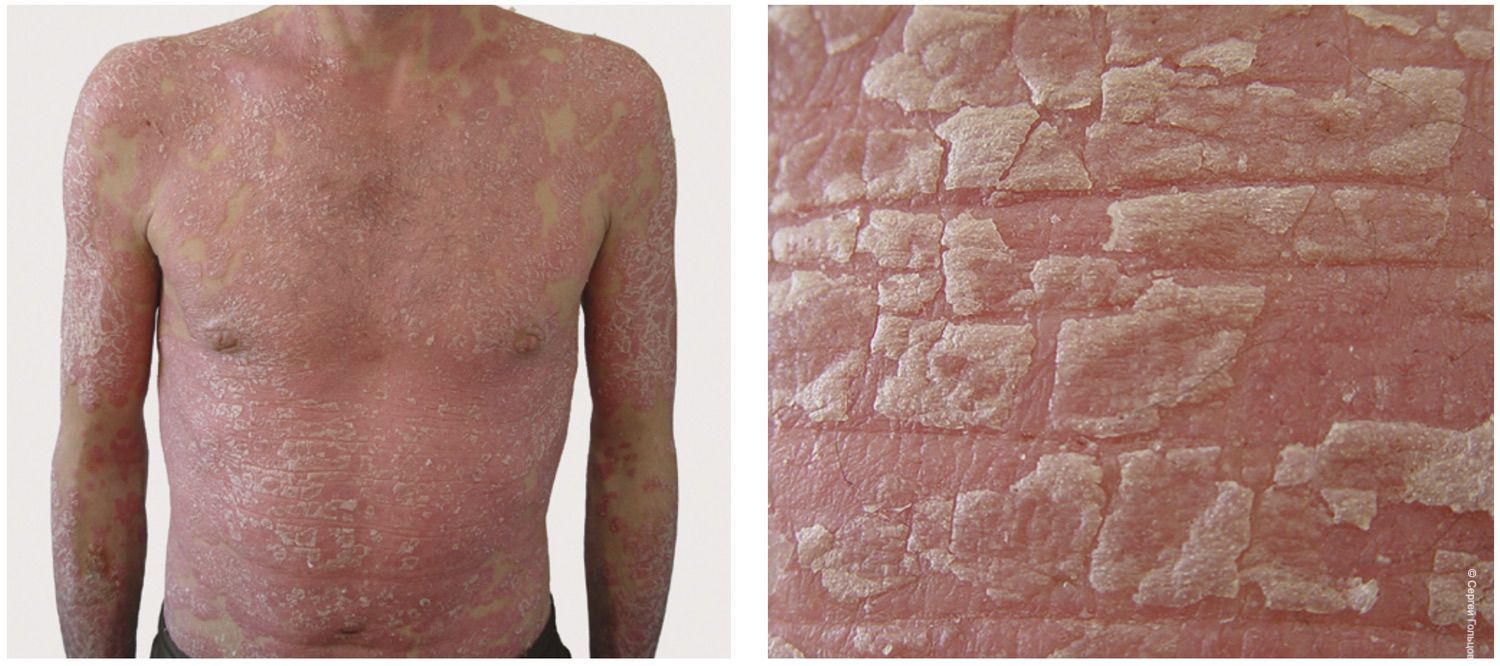
In our view, the treating physician needs to assess the cellular composition of the skin biopsy in order to document the cellular-level picture before treatment. How are the functions of each cell type expressed, and how did they generate the observed clinical picture? How are these functions expressed after treatment (Figure 42)
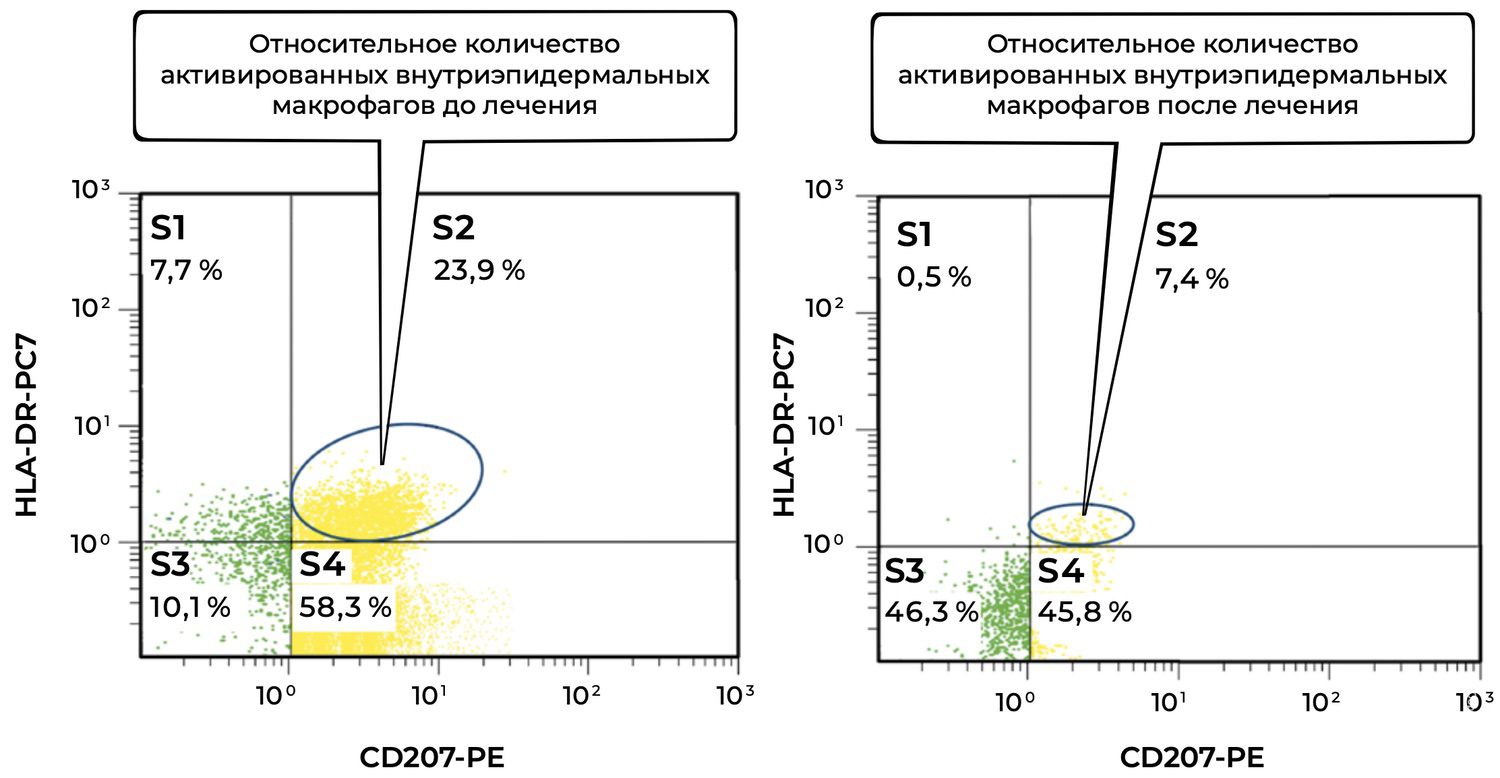
Figure 42. Comparative analysis of the relative proportion of viable activated intraepidermal macrophages (CD207⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of patient B., 51 years old, obtained on June 2, 2018 (No. 89/2), and June 17, 2018 (No. 89/3).
The cytoimmunogram shows the dynamics of the relative proportion of viable activated intraepidermal macrophages (CD207⁺ HLA-DR⁺ phenotype) in the skin-biopsy cell suspension of a patient with psoriasis before and after treatment.
Interpretation of Results
Before treatment:
The proportion of activated intraepidermal macrophages (CD207⁺ HLA-DR⁺) was 23.9%, reflecting pronounced local immune activation and participation of Langerhans cells in the pathogenesis of psoriasis.
The high proportion of these cells corresponds to immunopathological activity typical of psoriatic inflammation.
After treatment:
The relative proportion of activated macrophages decreased nearly threefold, to 7.4%, indicating reduction of the local immune response, suppression of inflammation, and restoration of cutaneous homeostasis.
At the same time, an increase in non-activated HLA-DR⁻ CD207⁺ cells was observed, along with a pronounced increase in the S3 population (CD207⁻ HLA-DR⁻), which may reflect a return toward the physiological state of the epidermal immune environment.
Overall, this patient demonstrated:
a substantial reduction in activation of CD207⁺ HLA-DR⁺ intraepidermal macrophages, demonstrating therapeutic efficacy;
restoration of epidermal cellular composition through an increase in viable but non-activated immune cells;
transition of the immune response from chronic inflammation toward remission.
These data indicate positive treatment dynamics, with suppression of the pathological immune inflammation characteristic of psoriasis. However, when the identified phenotype is correlated with the use of agents that act specifically on the detected parameters, the patient’s treatment becomes more targeted and object-directed.
If such agents have not yet been developed, the patient’s skin cytoimmunogram may nevertheless be used for objective evaluation of treatment efficacy and for documenting the state of the skin over time in the course of a chronic disease.
The chronic stage of eczema is characterized by infiltration and accentuation of the skin markings in the affected area, as well as post-inflammatory hypo- and hyperpigmentation. Persistent signs of eczema include pruritus, which intensifies during exacerbations, burning, and tenderness within the lesions (Figure 43). The symptoms described above characterize a rather broad group of skin diseases. Yet eczema accounts for up to 40% of the overall structure of cutaneous pathology. As in most skin diseases, histological examination of skin biopsy specimens is performed for differential diagnosis. However, it is not used for treatment selection, which is striking.
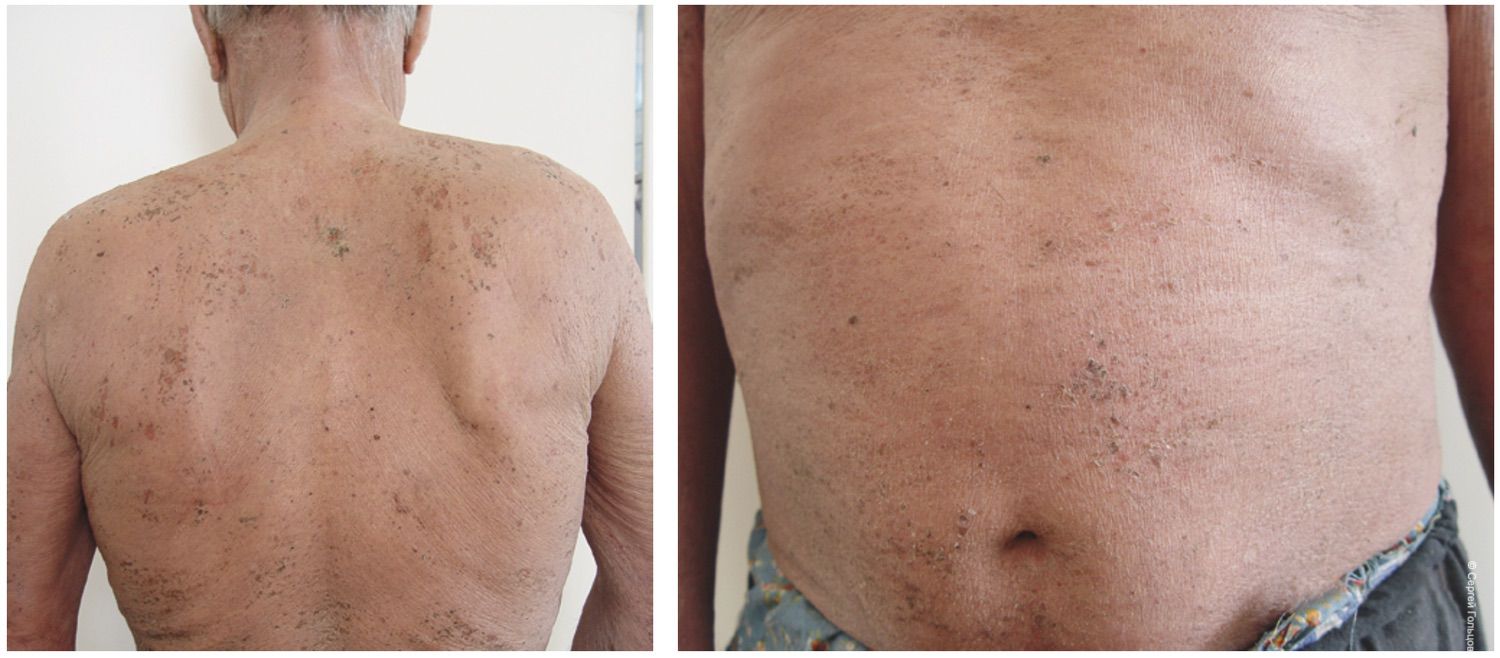
Figure 43. Clinical presentation of the local status of patient L., 48 years old, November 3, 2014, No. 39/2.
In the acute process, spongiosis is observed, with numerous small vesicles in the epidermis; intracellular edema is present in the cells of the spinous layer. In the dermis, dilation of the superficial vascular network, papillary edema, and lymphoid-cell infiltration around vessels are observed. In the chronic process, the dermis shows a perivascular infiltrate consisting of lymphocytes, fibroblasts, histiocytes, and eosinophils; the epidermis demonstrates acanthosis, hyperkeratosis, parakeratosis, and mild edema.
The skin cytoimmunogram makes it possible to add a functional characterization of skin cells to this description (Table 7), which is especially necessary for assessing the state of the skin as an actively functioning tissue in the age group that accounts for more than half of all affected patients (Figure 44).
Table 7. Skin Cytoimmunogram of Patient L., 48 Years Old, November 3, 2014, No. 39/2
|
Субпопуляции клеток кожи и жизнеспособность |
Фенотип |
Показатели, % |
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
66 3,5 |
|
Фибробласты, из них активированные |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
69 4,2 |
|
Клетки Лангерганса, из них активированные |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
45 1,0 5,0 0,3 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
1,0 0,1 19,7 1,7 1,5 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
75,4 8,9 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
4,5 1,7 |
|
Эпидермальные лимфоциты: Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
13 11 2 4 7 |
|
Жизнеспособность, % |
89 |
Cell viability was 89%, indicating a high-quality sample with preservation of cellular homeostasis. The skin phenotype demonstrates an inflammatory–regenerative type: a combination of moderate immune activity with signs of repair and stabilization of the tissue microenvironment. The cellular profile reflects the mixed Th2/Th1 response typical of chronic eczema, with moderate mast-cell degranulation, activation of keratinocytes and fibroblasts, and involvement of endothelial elements.
The analysis of cellular subpopulations is presented below.
Keratinocytes (CD49f⁺ HLA-DR⁺).
The total proportion was 66%, with activated forms accounting for 3.5%. Moderate epidermal activation was observed, reflecting a balance between inflammatory stimulation and repair processes. Pronounced HLA-DR hyperexpression was absent, distinguishing this phenotype from atopic dermatitis and psoriasis. The epidermis is functionally active but not hyperreactive; this state corresponds to remission.
Fibroblasts (CD45⁻ CD14⁻ CD44⁺ / CD80⁺).
The total proportion was 69%, with activated forms accounting for 4.2%. Increased activity of dermal fibroblasts was observed, indicating extracellular matrix remodeling and repair processes. This profile is typical of chronic eczema, in which inflammation is accompanied by a persistent attempt to restore the barrier structures of the skin.
Langerhans Cells (CD207⁺).
The total proportion was 45%, with activated phenotypes (CD80⁺ HLA-DR⁺/⁻) accounting for approximately 6%. Antigen-presenting activity was moderate. This parameter reflects reduced immune-surveillance intensity compared with the acute phase, while preserving a basal level of antigen recognition. The phenotype is typical of persistent subclinical inflammation.
Endothelial Cells (CD146⁺ CD54⁺ CD34⁺).
The cumulative proportion was approximately 23%, with predominance of CD54⁺ HLA-DR⁻ cells (approximately 19.7%). This is a sign of moderate endothelial activation and maintenance of the vascular component of inflammation. The presence of CD34⁺ cells (1.5%) indicates preserved angiogenic potential and microcirculatory adaptation.
Mast Cells (CD249⁺ CD63⁺).
The total proportion was 75.4%, with activated forms accounting for 8.9%, indicating high degranulation. This is an important phenotypic marker of eczematous processes: mast-cell activation initiates pruritus, erythema, and vascular reactivity. Mast-cell activity remains a key factor in chronicity.
Monocytes (CD45⁺ CD14⁺ HLA-DR⁺).
The total proportion was 4.5%, with activated forms accounting for 1.7%. The phagocytic compartment was moderately active, reflecting a balance between clearance and maintenance of inflammation. Reduced activity compared with acute forms is a sign of process stabilization.
Epidermal Lymphocytes:
Total T cells (CD3⁺): 13%;
T-helper cells (CD4⁺): 11%;
Cytotoxic T cells (CD8⁺): 2%;
B lymphocytes (CD19⁺): 4%;
NK cells (CD16⁺ CD56⁺): 7%.
The immune profile is Th2-dominant, with a moderate cytotoxic component. This distribution is characteristic of chronic eczematous states, in which IL-4/IL-13-dependent inflammatory mechanisms predominate, while innate control through NK cells is preserved.
The skin phenotype of patient L. indicates moderately active chronic inflammation with pronounced signs of regenerative compensation. Its principal features include:
preservation of epidermal barrier function with moderate HLA-DR activation;
marked participation of fibroblasts and endothelium in remodeling and angiogenesis;
a mast-cell component serving as a sustaining factor of inflammation;
a Th2-dominant immune profile with moderate cytotoxic involvement.
This type of skin cytoimmunogram corresponds to an inflammatory–reparative phenotype typical of chronic eczema. The skin cytoimmunogram of patient L. reflects stable chronic inflammation with signs of recovery:
moderate keratinocyte activation;
active fibroblasts and angiogenesis;
a significant mast-cell component;
a Th2-dominant profile;
low antigen presentation.
Thus, the skin is in a phenotypic state of “remission inflammation”—a balance between inflammation and repair. This phenotype is favorable for targeted regenerative therapy, including agents that enhance trophic support and normalize the cutaneous microenvironment.
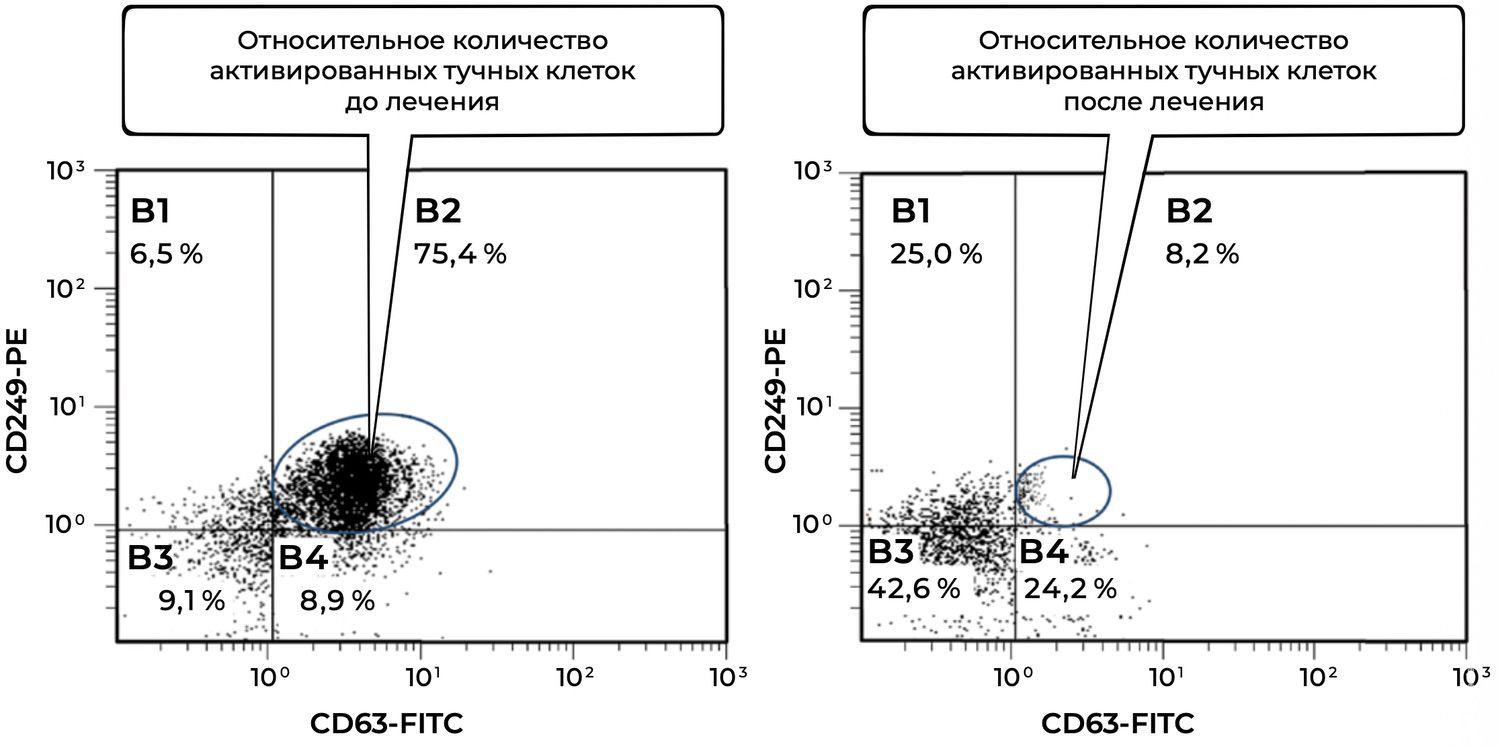
Figure 44. Comparative analysis of the relative proportion of viable activated mast cells (CD249⁺ CD63⁺ phenotype) in the skin-biopsy cell suspension of patient L., 48 years old, obtained on November 3, 2014 (No. 39/2), and November 18, 2014 (No. 39/3).
The presented pair of cytoimmunograms shows flow-cytometric analysis of the relative proportion of viable activated mast cells (CD249⁺ CD63⁺ phenotype) in skin-biopsy cell suspensions from a patient with chronic eczema before and after treatment.
Interpretation of Results
Before treatment:
An extremely high proportion of activated mast cells was observed: 75.4%. This reflects a pronounced allergic and inflammatory background characteristic of chronic eczema.
The proportion of non-activated mast cells (CD249⁺ CD63⁻) was only 8.9%, indicating widespread mast-cell activation in the skin.
After treatment:
The proportion of activated mast cells decreased sharply, almost tenfold, from 75.4% to 8.2%, indicating effective suppression of inflammation.
The population of viable non-activated mast cells (CD249⁺ CD63⁻) increased to 24.2%, and the CD249⁻ CD63⁻ population (B3) increased to 42.6%. This may indicate restoration of the cutaneous immune balance and possible repopulation of the cellular pool.
In conclusion, in this patient with chronic eczema:
marked mast-cell hyperactivation was initially present, involving the key mediators of allergic and inflammatory skin reactions;
after therapy, a pronounced reduction in mast-cell activation occurred, corresponding to clinical remission and suppression of the inflammatory cascade;
restoration of the proportion of viable non-activated cells indicates preservation of the tissue reserve and the potential for a normal immune response.
The diagrams convincingly demonstrate the immunomodulatory effect of therapy and its high effectiveness in the treatment of chronic eczema.
The examples presented demonstrate the effectiveness of obtaining skin cytoimmunograms for assessing skin status before treatment, at the achievement of clinical remission, and as criteria for evaluating therapeutic efficacy. Precision diagnosis—more accurate because it is expressed quantitatively—already establishes a foundation for future practical applications.
However, there are far more reasons to develop interventions with predefined cellular phenotype parameters.
Any injury to the skin initiates a complex cascade of temporally and spatially organized events that ensure restoration of homeostasis and closure of the wound surface. This process comprises several phases—inflammation, proliferation (granulation tissue formation and re-epithelialization), and remodeling—each involving specific cellular and molecular mechanisms. At every stage, these processes are regulated by soluble factors that possess distinct regulatory functions and therefore provide opportunities for therapeutic intervention. This served as the rationale for the development of a biotechnological product with the intended effect of promoting wound healing.
Skin cells occupy specific positions within tissue architecture and derive functional significance from these locations. Epithelial stem cells situated within the hair follicle niche serve as progenitors of cellular populations responsible for the formation of epithelial skin structures, including the interfollicular epidermis, hair follicles, sebaceous glands, sweat glands, and related cellular lineages. Following injury, they become a source of interfollicular epithelial regeneration and exert local anti-inflammatory effects by restoring the balance between pro- and anti-inflammatory cytokines within damaged tissue.
Given that granulation tissue formation is accompanied by the release of a broad spectrum of growth factors and cytokines stimulating keratinocyte proliferation, re-epithelialization might appear not to be a major challenge. In reality, however, deep skin injuries destroy not only the epidermis but also epithelial appendages such as hair follicles and glands. This disrupts the mechanisms of re-epithelialization and dermal histoarchitecture, which depend upon the structural micro-modules formed by follicles and glands. Complete restoration of epidermal and epithelial skin structures is therefore possible only if the pool of epithelial stem cells is preserved.
The understanding of epithelial stem-cell biology expanded substantially following the discovery of a relatively small population of endothelial progenitor cells within the CD34⁺ hematopoietic cell fraction of bone marrow mononuclear cells. During vascular remodeling in damaged tissues, which involves collateral arteriole development and capillary branching, de novo microvessel formation depends on endothelial progenitor cells that migrate from the bone marrow into the peripheral circulation and subsequently to ischemic sites.
It has been demonstrated that ischemia increases the number of these cells, and evidence exists that they can incorporate into capillary walls and subsequently differentiate into mature endothelial cells. Without the capacity of endothelial cells to restore vascular networks and support tissue growth, skin repair would be impossible. This alone identifies endothelial cells as key regulators of the reparative process, since both their abundance and functional activity determine the outcome of tissue repair. Moreover, endothelial progenitor cells, similarly to other CD34⁺ hematopoietic cells, are capable of activating resident endothelial cells through angiogenic factors.
Loss of epithelial stem cells and endothelial progenitor cells results in a progressive deficit of reparative potential, which inevitably manifests during the subsequent remodeling phase of wound healing. Remodeling involves the restructuring of granulation tissue. If cellular and molecular deficits in reparative capacity are not compensated in a timely manner, granulation tissue is replaced by mature connective tissue, leading to scar formation.
To compensate for deficits in reparative potential and stimulate healing of extensive skin wounds, the introduction of cell-based technologies into clinical practice has resulted in widespread use of mesenchymal stromal stem cells derived from bone marrow and autologous fibroblasts isolated from the dermis. These developments have also reinforced the concept of the “stem-cell niche.”
The concept of a niche encompasses the cellular microenvironment and extracellular matrix specific to stem cells and capable of regulating their fate. The niche produces chemical factors controlling cellular proliferation, selection, and differentiation. According to current understanding, the stem-cell niche constitutes a specialized microsphere that includes both cellular components and surrounding matrix architecture. This environment preserves stem-cell populations and regulates the stages of their differentiation. One group of cells undergoes post-mitotic apoptosis at an early stage; a second group, resistant to apoptosis and differentiation, participates in niche formation; and a third group consists of immature cells that mature into fully differentiated cells.
The niche of epidermal epithelial stem cells is spatially restricted. Each niche contains a cluster of proliferating and differentiating cells. At the center of each group of basal cells is a single stem cell. Through asymmetric division and approximately four rounds of proliferation, this stem cell generates a clone of 9–11 basal cells. Stem-cell properties are maintained through contact with the basement membrane, which itself functions as a niche. Loss of this interaction leads to the formation of a transit-amplifying pool followed by terminal differentiation. These cellular groups are arranged as hexagonal units whose progeny migrate upward into the suprabasal layers and eventually become cornified keratinocytes.
Thus, reparative potential at the cellular level may be summarized as follows:
tissue renewal is maintained by continuous proliferation and differentiation of stem cells located along the epidermal basement membrane and the hair-follicle interface;
basal epidermal stem cells sustain the keratinocyte pool;
hair-follicle cells are multipotent and capable of differentiating into keratinocytes, fibroblasts, and other dermal cell types;
the principal function of stem cells is physiological replacement of senescent differentiated skin cells and restoration of cellular reserves.
Activation of stem cells requires cytokines, growth factors, and other signaling molecules. Their synthesis is regulated both by the stem cells themselves and by their microenvironment—the niche—which provides signals determining whether stem cells maintain their identity or differentiate into specialized mature cells.
With aging, in accordance with the Hayflick limit, the total number of stem cells participating in tissue repair declines, particularly mesenchymal stem cells. Cellular depletion and reduced synthetic activity lead to deficiencies of cytokines, growth factors, signaling molecules, and regulatory mediators, resulting in diminished proliferative activity. The importance of signaling molecules in mediating cellular interactions thus becomes increasingly evident.
The ability of cells to perceive and respond to signals is a fundamental property of life. In multicellular organisms, cells with diverse functions exchange a wide variety of signaling molecules. The regulatory role of these molecules, and the timely compensation of their age-related or post-traumatic deficiency, appears to offer promising opportunities for optimizing topical therapy of skin injuries and disorders. Experimental studies have already demonstrated the effectiveness of such approaches.
A group of investigators analyzed the gelatinous embryonic substance of the umbilical cord, a source of embryonic stem cells, peptides, signaling molecules, growth factors, phospholipids, enzymes, and glycosaminoglycans. Among the molecules identified was Wharton Jelly Peptide P199, a polypeptide capable of stimulating stem-cell division. Subsequent studies investigated its effects on the synthesis of signaling molecules required for activation of skin stem-cell proliferation. This polypeptide was shown to significantly increase cytokine and growth-factor production in cultured human skin cells, thereby generating a cascade of signaling molecules necessary for proliferation and differentiation. However, the specific signaling molecules responsible for intercellular communication and subsequent cell division were not identified.
Another important feature of cellular signal transduction is signal amplification. The ultimate cellular response depends on the number of effector molecules—enzymes, structural proteins, transporters, and others—that are influenced by the signal. Direct interaction between signaling molecules and effectors would require enormous quantities of signaling factors. To overcome this limitation, biological systems employ amplification mechanisms involving secondary messengers.
Equally important is signal termination. Signal-transduction pathways incorporate feedback mechanisms that deactivate receptors, remove them from the cell surface through endocytosis, or interrupt downstream signaling through regulatory proteins such as G proteins. Another critical feature is signal integration—the ability of a cell to receive multiple diverse signals and generate a coordinated response appropriate to its physiological requirements.
Different signaling pathways intersect at multiple levels, creating a complex network of interactions that maintain cellular and organismal homeostasis under both physiological conditions and in response to skin injury.
Since wound healing depends on replenishing deficits of cells and signaling molecules within damaged tissue, the concept of introducing an external initiating signal into a wound—compensating for age-related and post-traumatic deficiencies and thereby stimulating proliferation, amplification of progenitor-cell populations, and enhanced reparative function—appears both rational and promising. Consequently, the search for a specific signaling molecule capable of initiating such proliferative activity is justified.
It is not coincidental that, under the requirements of Federal Law No. 180-FZ “On Biomedical Cell Products,” all newly developed cell-containing products must be standardized, and flow cytometry has been established as the principal method for quality assessment of biomedical cellular products.
A distinct form of precision therapy involves the use of biomedical cellular products, their derivatives, and medical devices based upon them. Although several such products are already approved for clinical use worldwide, most are based on allogeneic human cells. Personalized approaches utilize autologous cells, in which donor and recipient are the same individual. However, cells and biologically active molecules may also be obtained from tissues of other animal species and used for diverse therapeutic purposes.
Advances in molecular biology have enabled the industrial production of biologically active proteins identical to endogenous human molecules. This methodology relies on isolation or synthetic generation of DNA encoding the desired protein, transfer into microbial or mammalian producer cells, induction of hypersecretion, and large-scale expansion of cell cultures. These approaches ultimately gave rise to the scientific and industrial discipline of biotechnology.
Cell cultures provide the principal platform for monitoring proliferation, assessing viability, morphology, and differentiation, studying responses to stimuli, analyzing intercellular interactions, evaluating migration into surrounding tissues, stimulating angiogenesis, and investigating production of biologically active molecules for novel therapeutic applications.
Research on intercellular interactions dates back to the work of A. Moscona on selective self-aggregation of embryonic cells. These studies were extended by M. Steinberg, whose theory of differential adhesion explained organ and tissue morphogenesis. Investigations of epithelial–mesenchymal plasticity demonstrated the feasibility of creating three-dimensional systems in which cytodifferentiation and extracellular matrix influences were minimized. In spheroid cultures, cells restored tissue organization through selective intercellular interactions, demonstrating a “universal response to injury” and revealing unprecedented opportunities for applying xenogeneic embryonic systems to tissue repair.
The term xenogeneic (from the Greek xenos, foreign, and genos, origin) refers to tissue used to compensate for tissue deficits in humans that is derived not from another person or artificially engineered constructs but from donor animals of another biological species. Such biological materials differ from synthetic alternatives and may avoid many associated adverse effects.
The possibility of using xenogeneic biological sources for the creation of biotechnological products has encouraged the search for novel sources of stem cells and their signaling molecules. Animal embryonic tissues represent one of the most accessible and renewable sources of such materials.
Several significant advantages have been identified for chicken embryos, which are regarded not only as a safe source of donor cells for vaccine production but also as a means of obtaining cells in virtually unlimited quantities under a quality-control system that has existed for decades. The advantages of chicken embryonic cells become particularly evident when compared with the use of human fetal tissue, which is associated with numerous challenges, including ethical concerns, insufficient donor control, determination of the optimal gestational age for cell isolation, assessment of donor-tissue quality, limited scientific understanding, and legal restrictions on the use of human fetal tissues. Chicken embryonic cells can be harvested at precisely defined stages of gestation, making it possible to standardize the quality and viability of the producer cells required for obtaining biologically active molecules.
Another obstacle to the successful application of xenogeneic cells is graft rejection. Cell transplantation does not eliminate problems associated with the immune rejection of foreign tissues. Alongside the beneficial effects of cellular transplantation, immunogenic responses should also be anticipated. This issue, however, is entirely avoided when molecules isolated from cellular homogenates are transferred in the form of a supernatant. The ability to obtain biologically active molecules in soluble form distinguishes molecular therapy from cellular therapy, since it does not induce undesirable immune recognition of donor-cell membrane proteins.
A method used to isolate and precipitate a target molecule from a mixture through the formation of an antigen–antibody complex is known as immunoprecipitation. Expanding our understanding of the immunopathogenic mechanisms of wound healing may ultimately enable therapeutic approaches aimed at compensating for deficiencies in a patient’s own cells, active molecules, or even tissues. However, xenogeneic transfer requires a reliable source of biological material.
Thus, the objective became both precise and clearly defined:
To develop a biotechnological product for enhancing skin repair in vivo, containing as its active ingredient adhesion molecules derived from xenogeneic cells expressing the CD34⁺CD45dim phenotype.
According to Article 1 of EU Regulation No. 2001/393/EC, an SPF embryo is free of specific pathogenic contaminants, including bacteria, fungi, viruses, and antibodies directed against these microorganisms. Furthermore, Article 2(2) of Council Directive 90/539/EEC states that SPF embryos may be obtained from birds certified as free of specific pathogens under the European Pharmacopoeia and are intended exclusively for diagnostic, research, or pharmaceutical purposes. Since February 1, 2023, Russia has also implemented a national standard governing incubation eggs used as biological raw materials. According to Veronika Skvortsova, Head of the Federal Medical-Biological Agency of Russia:
“The FMBA of Russia acted as the expert organization during development of this standard because the Agency possesses extensive experience using incubation eggs as raw materials for influenza vaccine production. Rosstandart approved the standard on December 29, 2022. Strict requirements for raw materials are essential because sterile conditions must be ensured for both poultry maintenance and egg production. The use of embryos obtained from SPF-certified eggs reduces contamination risks to virtually zero and opens significant opportunities for production and export.”
Consequently, the principal stage in developing the biotechnological wound-healing product involved the isolation of a xenogeneic active substance.
To achieve this, sterile SPF chicken embryos at six days of gestation were placed into 2-mL cryovials containing a freezing medium composed of 90% fetal bovine serum (FBS) and 10% DMSO as a cryoprotectant. Samples were cooled in liquid-nitrogen vapor at a rate of 1°C per minute to −40°C and subsequently transferred into liquid nitrogen (−170°C) for long-term storage in Dewar vessels.
For preparation of cellular suspensions, frozen material was thawed ex tempore in nitrogen vapor to preserve maximal cell viability. The material was transferred into sterile glass centrifuge tubes, washed with sterile 0.9% sodium chloride solution to remove the freezing medium, and centrifuged for 10 minutes at 1,500 rpm. The supernatant was removed, and the washing procedure was repeated. Three milliliters of 0.9% sodium chloride solution were added to the pellet and homogenized, followed by an additional 5 mL of saline.
To sediment stromal components, centrifugation was performed at 1,500 rpm: brain-tissue cells for 1 minute and liver and other tissue cells for 10 seconds. The resulting supernatant was transferred into sterile tubes and centrifuged again for 5 minutes at 1,500 rpm. The supernatant was discarded, and 3 mL of 0.9% sodium chloride solution were added to the pellet. Following resuspension, the volume was adjusted to 10 mL with saline and centrifuged again for 5 minutes at 1,500 rpm.
The resulting suspension was analyzed for cellular phenotype, viability, and total cell count. Viability of CD34⁺CD45dim cells was required to be at least 80%; samples below this threshold were discarded. The cell concentration was then adjusted with 0.9% sodium chloride solution to 1.0 × 10⁶ ± 0.25 × 10⁶ cells/mL.
The present invention pertains to physiology, medicine, surgery, traumatology, and clinical pharmacology and may be used for manufacturing a biotechnological product designed to accelerate healing of wounds of various etiologies. Before clinical application, however, the effects of its active substance on processes directly involved in tissue repair had to be evaluated.
One such process is apoptosis, which plays a critical role in wound healing. If apoptosis does not occur appropriately, inflammatory processes persist because necrotic cells continue releasing pro-inflammatory mediators. Conversely, delayed apoptosis may result in replacement of granulation tissue by mature scar tissue. Understanding the role of apoptosis in scar formation therefore offers opportunities for controlling repair processes through modulation of immune mechanisms governing cell death.
To investigate this phenomenon, camptothecin, a known inducer of apoptosis through inhibition of DNA topoisomerase I activity, was used at final concentrations of 1.0 and 0.2 μM. The xenogeneic test substance was applied at final concentrations of 7, 70, and 350 μg/mL.
THP-1 suspension cultures were maintained in RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 50 μg/mL gentamicin, and 2 mM L-glutamine. Fetal bovine serum remains the gold-standard culture supplement because it provides nutritional and hormonal support as well as binding proteins necessary for cellular proliferation, differentiation, and adhesion.
Cells were incubated with both stimulators simultaneously for 24 hours at 37°C in a humidified atmosphere containing 5% CO₂. Cell passages were performed every 2–3 days. Cultures were maintained in 50-mL plastic flasks (Sarstedt, Germany). For experimental procedures, 200 μL of cell suspension (2 × 10⁶ cells/mL) were added to wells of 96-well flat-bottom plates.
To determine the proportion of cells at different stages of apoptosis, two fluorescent dyes—YO-PRO-1 and propidium iodide (PI)—were employed. One hundred microliters of cell suspension (2–3 × 10⁶ cells/mL) were stained with YO-PRO-1 at a final concentration of 250 nM and PI at 1 μg/mL. Samples were incubated for 15 minutes at room temperature in the dark, diluted with phosphate-buffered saline, and analyzed using a Navios flow cytometer (Beckman Coulter, USA). At least 20,000 single cells were analyzed per sample.
The concentrations of camptothecin and the xenogeneic substance were denoted as coordinate sets (xᵢ, yᵢ), with corresponding percentages of cells undergoing early apoptosis (zᵢ) and late apoptosis/necrosis (z'ᵢ). These datasets generated two matrices representing points on separate three-dimensional surfaces. Data processing was performed using Python with the SciPy and NumPy libraries. Two-dimensional mesh grids were created using numpy.meshgrid, and interpolation was performed using scipy.interpolate.griddata. Surface visualization was generated using Matplotlib. Results were expressed as percentages of positive cells.
Flow-cytometric data were processed using EXPO-32, CXP v2.2, Navios Software v1.2, Kaluza™ v1.2 (Beckman Coulter), and Statistica 8.0 (StatSoft).
In control THP-1 samples, viable cells accounted for 96.53 ± 0.46%. Exposure to the xenogeneic substance at 350 μg/mL significantly reduced viability (p < 0.001) to 91.72 ± 0.44%. Simultaneously, the proportion of cells in early apoptosis increased significantly (3.16 ± 0.19% vs. 1.99 ± 0.39%, p = 0.024), while late apoptotic/necrotic cells increased more than fourfold (5.12 ± 0.28% vs. 1.47 ± 0.21%, p < 0.001).
At 70 μg/mL, the substance produced a significant twofold increase (p = 0.009) in terminal apoptotic and necrotic cells (2.69 ± 0.29%). At 7 μg/mL, no significant effects were observed.
Exposure to 1 μM camptothecin dramatically reduced viability to 31.58 ± 1.68% (p < 0.001) and increased the proportion of cells in early and late apoptosis to 30.12 ± 1.94% and 38.30%, respectively. However, in the presence of the xenogeneic substance at 350 μg/mL, viability increased significantly to 45.11 ± 2.17% (p = 0.001), while early apoptotic cells decreased to 21.49 ± 2.15% (p = 0.018) and late apoptotic/necrotic cells decreased to 33.40 ± 2.49%.
Lower concentrations of the test substance (70 and 7 μg/mL) did not significantly alter apoptotic distributions.
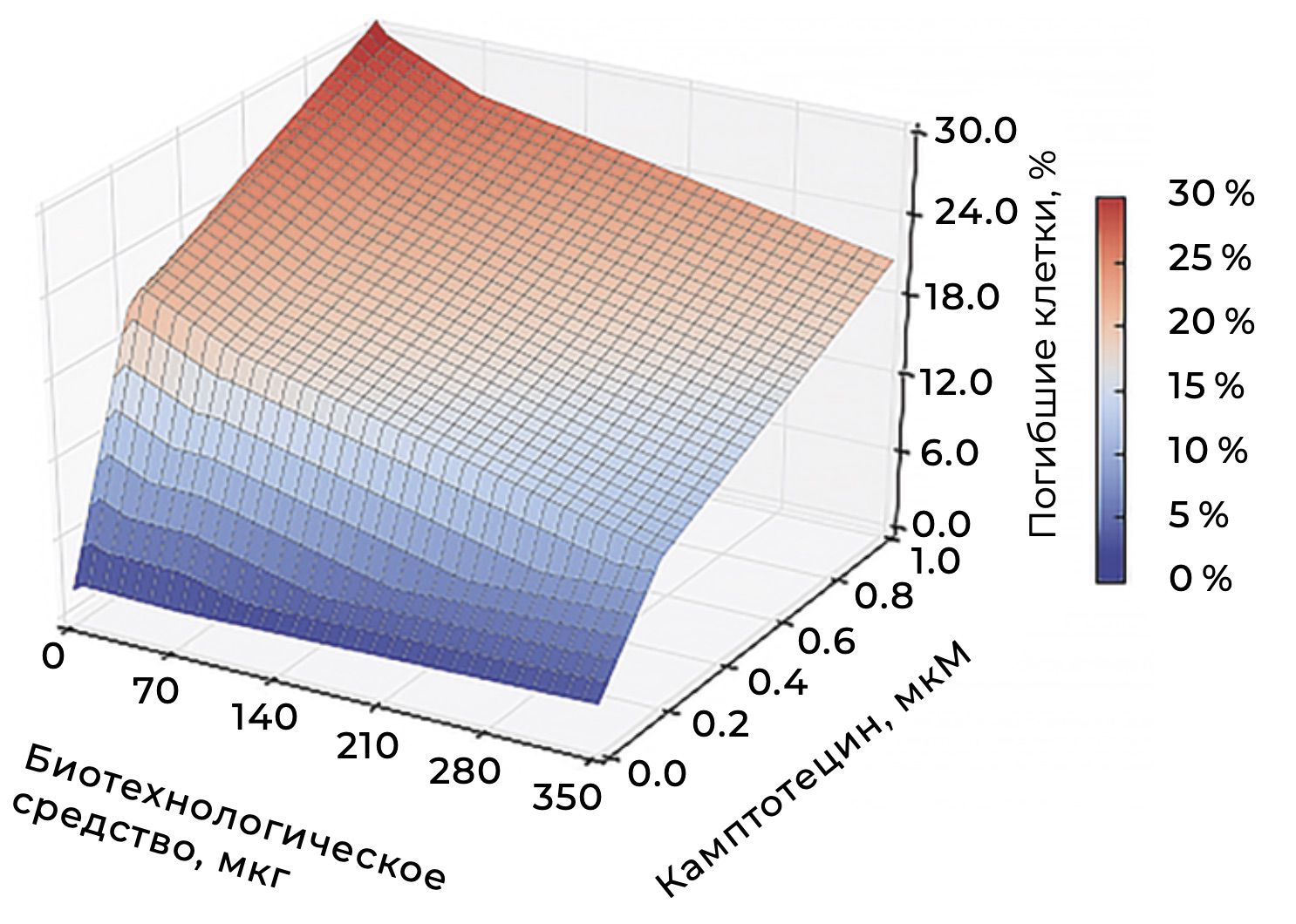
Figure 45 illustrates the dependence of early apoptosis on concentrations of both compounds. The color gradient ranges from blue (minimal apoptosis) to red (maximal apoptosis). The graph demonstrates that the biotechnological substance alone exhibits low cytotoxicity, whereas camptothecin induces dose-dependent apoptosis. Combined exposure produces enhanced apoptotic responses, with maximal cell death observed at high concentrations of both agents.
Figure 46 depicts late apoptosis and necrosis. Here, the color gradient ranges from brown (low cell death) to violet (high cell death). The biotechnological substance alone does not induce substantial late cell death. Camptothecin, however, causes pronounced dose-dependent cytotoxicity, with combined exposure producing the highest levels of cell death.
When the concentration of camptothecin was reduced to 0.2 μM, viable-cell content increased to 47.76 ± 3.17%, while early and late apoptotic cells accounted for 20.75 ± 2.40% and 31.49 ± 1.31%, respectively. Under these conditions, co-incubation with 350 μg/mL of the biotechnological substance did not significantly affect overall viability (51.11 ± 1.49%, p = 0.367) but nearly halved the proportion of cells in early apoptosis (12.86 ± 2.00%, p = 0.036).
These findings indicate a substantial reduction in overall cytotoxicity when camptothecin is used at a minimally effective concentration. Although apoptotic processes remain active, lowering the dose preserves a significant proportion of viable cells and may therefore improve the therapeutic balance between efficacy and safety.
The principal conclusions were as follows:
reducing camptothecin concentration to 0.2 μM significantly improves cell survival;
apoptosis remains present but is less pronounced and progresses more slowly;
lower concentrations reduce the likelihood of necrosis-associated inflammation and tissue injury;
combining camptothecin with the biotechnological substance provides a more favorable balance between biological activity and safety.
Overall, the study demonstrated a dose-dependent influence of the isolated xenogeneic active substance on cell viability and apoptotic processes in culture. The resulting data suggest that the reparative process can be modulated experimentally, thereby providing a rationale for subsequent wound-healing studies in laboratory animals.
The experimental phase of the study involved 80 adult male Chinchilla rabbits obtained from a breeding facility in Tyumen. Animal housing and all experimental procedures were conducted in accordance with the regulations of the Council of the European Communities (Directive 86/609/EEC, November 24, 1986). Specifically, the animals were maintained individually in standard vivarium cages under identical lighting and temperature conditions and received a conventional diet according to established husbandry guidelines. The study was performed during the autumn–winter season. At the conclusion of the experiment, animals were euthanized under ether anesthesia in accordance with accepted ethical standards.
Experimental full-thickness skin wounds were created using a standardized protocol in all animals. An infected soft-tissue wound model (Russian Patent No. 2006122640) was employed. All procedures were performed under ether anesthesia.
The dorsal interscapular region along the paravertebral line was clipped and carefully shaved using a safety razor. A circular metal template with an internal diameter of 2.0 cm was then used to excise a full-thickness skin segment measuring approximately 250 mm² (Figure 47).
Figure 47. Preparation of a laboratory animal for induction of an infected soft-tissue wound model.
The resulting defect was treated with 70% acetic acid until necrosis of the subcutaneous tissue and underlying muscle was achieved (Figure 48).
Figure 48. Appearance of wounds following treatment with 70% acetic acid, Day 1 of the experiment.
Subsequently, 500,000 CFU of Staphylococcus aureus were introduced into the wound. After 48 hours, by the start of the treatment phase, all animals had developed infected wounds of irregular circular shape measuring up to 200 mm² (Figure 49).
Figure 49. Induction of an infected soft-tissue wound model.
An inflammatory infiltrate measuring up to 300 mm² formed around the wound. The wound margins were hyperemic, firm, and adherent to surrounding tissues. The wound bed was covered with a dense fibrin layer and produced approximately 2–3 mL of purulent exudate of whitish color and paste-like consistency.
On the day the wound was established, a full-thickness biopsy extending to a depth of 1.8–2.2 cm was obtained under local anesthesia. Tissue samples were fixed in 96% ethanol, embedded in paraffin, sectioned, and stained with Mayer’s hematoxylin and eosin for subsequent histological evaluation.
Prior to treatment, histological examination in all animals revealed foci of necrosis within connective and muscular tissues (Figure 50), together with vascular dilation, blood stasis, and leukocyte exudation.
Figure 50. Histological characteristics of wounds before treatment. Mayer’s hematoxylin and eosin staining, ×40 magnification. Foci of necrosis within connective and muscle tissues.
The papillary dermis demonstrated distinct stratification with pronounced cellular infiltration consisting primarily of monocytes, occasional macrophages, and loose connective tissue containing numerous fibrocytes, isolated fibroblasts, monocytes, and plasma cells. Areas of necrosis were replaced predominantly by polymorphonuclear leukocytes, accompanied by mononuclear lymphocytes (Figure 51).
Figure 51. Histological characteristics of wounds before treatment. Mayer’s hematoxylin and eosin staining, ×40 magnification.
Treatment was initiated five days after induction of the infected wound model. The animals were randomly assigned to four groups, each consisting of 20 rabbits. Wounds were managed using an open-healing approach.
Dressings were performed once daily according to the treatment regimen assigned to each group:
Group I (Experimental Group).
The wound received the active substance of the original formulation. After application, the gel was allowed to dry and form a film over 3–5 minutes, after which a sterile dry dressing was applied.
Group II (Vehicle Control).
The wound received only the gel base composed of hydroxyethylcellulose.
Group III (Conventional Therapy).
Treatment was performed according to standard wound-care principles. During the inflammatory phase, water-based ointments were used; during the regeneration and epithelialization phases, lipid-based ointments were applied.
Group IV (Untreated Control).
Following routine wound cleansing, only a sterile dry dressing was applied.
During visual observation of wound healing, several differences in overall condition and behavior were noted among the groups. A characteristic feature of animals in the experimental group was increased motor activity, greater food consumption, and improved coat condition.
Wound size, edema, hyperemia, wound exudate, and the nature of wound discharge were assessed on Days 1, 3, 5, 7, 10, and 15.
In all control groups, wound dimensions remained unchanged during the first day. Wound margins remained dense and adherent to surrounding tissues. A compact inflammatory infiltrate persisted around the wound. Mechanical pressure elicited purulent discharge, and the wound surface remained covered by a purulent film (Figure 52).
Figure 52. Appearance of infected soft-tissue wounds on Day 1 of the experiment.
By contrast, on Day 1 in Group I (Experimental Group), the wound area measured approximately 150 mm². Moderate hyperemia was present around the wound, the wound margins were soft on palpation, the wound bed was covered with fibrin deposits, and no purulent exudate was observed (Figure 53).
Figure 53. Appearance of infected soft-tissue wounds in Group I (Experimental Group). Day 1 of treatment.
By Day 3, the tissue defect had decreased to approximately 60 mm². The wound surface was covered by a pale gray scab, and a marginal epithelialization zone measuring up to 5 mm in width was observed around the wound (Figure 54).
Figure 54. Appearance of infected soft-tissue wounds in Group I (Experimental Group). Day 3 of the experiment.
By Day 5, wounds in Group I demonstrated closure of more than 70% of the original defect, accompanied by active marginal epithelialization and visible hair regrowth (Figure 55).
Figure 55. Appearance of infected soft-tissue wounds in Group I animals. Day 5 of the experiment.
In the experimental Group I, complete epithelialization of the wound defect was observed by Day 10, accompanied by restoration of hair growth and a histological pattern consistent with regeneration of all functional skin structures. Wound closure occurred without formation of connective-tissue scar tissue, and restoration of function in the damaged area was confirmed by regrowth of the hair coat (Figure 56).
Figure 56. Appearance of infected soft-tissue wounds in animals of Experimental Group I. Day 10 of the experiment.
In Group II (vehicle control), wounds largely retained their original dimensions throughout the first ten days of observation. Histological examination revealed edema, lymphohistiocytic infiltration, and degenerative-dystrophic changes involving the subcutaneous tissue and underlying muscle layer (Figure 57). Wound closure did not occur. As demonstrated in the photographs, only a slight reduction in wound size was observed during this period (180–200 mm²). The wound surface remained covered with fibrin deposits, while the wound edges and walls remained dense and indurated.
Figure 57. Appearance of infected soft-tissue wounds in Group II animals (vehicle control). Days 10 and 25 of the experiment.
In Group III (conventional treatment), all classical stages of wound healing were observed, including inflammation, proliferation, and regeneration (Figure 58).
Figure 58. Appearance of infected soft-tissue wounds in Group III animals (conventional treatment). Day 20 of the experiment.
Complete closure of the wound defect occurred by Day 25, accompanied by formation of a dense connective-tissue scar (Figure 59).
Figure 59. Appearance of infected soft-tissue wounds in Group III animals (conventional treatment). Day 25 of the experiment.
In Group IV, five animals developed subcutaneous phlegmon by Day 5, resulting in death (Figure 60).
Figure 60. Appearance of infected soft-tissue wounds in Group IV animals on Day 25 of the experiment. The wounds retained their original dimensions, the wound bed was covered with necrotic tissue, and abundant purulent discharge was present.
In the remaining animals of Group IV, wound closure was not achieved by Day 25, and the animals were subsequently removed from the experiment.
In the experimental group (Group I), application of the active xenogeneic substance resulted in clearance of purulent-necrotic tissue within 1–3 days of treatment, while epithelialization was completed by Days 5–7, accompanied by restoration of hair growth. In the control groups, wound cleansing occurred substantially later, at approximately Days 15–20 in Group II (vehicle control), whereas closure of the wound defect with formation of a dense connective-tissue scar occurred between Days 20 and 30 in Group III (conventional treatment) (Figure 61).
Figure 61. Visual comparison of wound-healing dynamics in infected soft-tissue wounds. Group III (conventional treatment, upper row) and Group I (experimental treatment, lower row).
To quantify wound-healing rates, a modified version of the L. N. Popova wound-healing test was employed. A sterile cellophane sheet was placed over the wound, and its outline was traced using ink. The tracing was then transferred onto graph paper, and the wound area was calculated by counting enclosed square millimeters. During subsequent measurements, the procedure was repeated, and the percentage reduction in wound area per day was calculated according to the formula:
[
\text{Healing Rate (%)} = \frac{(S - S_n)}{S \times t} \times 100
]
where:
S = wound area at the previous measurement;
Sₙ = wound area at the current measurement;
t = number of days between measurements.
Table 8. Comparative Characteristics of Wound Area During Sequential Stages of Healing
The results demonstrated pronounced differences among the treatment groups.
As early as Day 1, wound area in Group I decreased to 156 ± 2.5 mm², whereas wounds in the remaining groups measured approximately 218–245 mm². This difference was statistically significant.
Between Days 2 and 6, wound area in Group I further decreased to 38 ± 1.9 mm², while values in Groups II and III remained between 169 and 218 mm², and wounds in Group IV continued to enlarge. These differences were likewise statistically significant.
In Group III, epithelialization and scar formation occurred between Days 20 and 25, whereas in Group I wound healing was essentially complete by Day 7.
In Group II, wound area remained nearly unchanged (214–228 mm²) until approximately Day 13, significantly differing from the healing dynamics observed in Groups I and III.
In Group IV, partial data were obtained before removal of the animals from the experiment. Nevertheless, the trend was opposite to that observed in the treated groups, with wound areas increasing to 325–330 mm², a difference that was also statistically significant.
In summary, statistically significant differences (p < 0.05) were observed:
between Group I and all other groups beginning on the first day of treatment;
between Group III and Group II beginning on Days 3–4, reflecting faster healing in the conventionally treated group compared with vehicle control;
between Group IV and all other groups at nearly every observation point prior to animal withdrawal, due to progressive wound enlargement rather than contraction.
These findings indicate that the xenogeneic active substance exerted a pronounced stimulatory effect on tissue repair. Visual observations were subsequently correlated with histological changes.
Histological examination performed on Day 3 in animals of the experimental group revealed loose connective tissue containing leukocytic infiltrates, isolated enlarged fibroblasts, lymphocytes, plasma cells, eosinophils, and monocytes. Areas of endothelial proliferation were observed, characterized by immature vascular structures lacking lumina and exhibiting multilayered nuclear arrangements (Figure 62).
Figure 62. Histological characteristics of infected wounds in animals of the experimental group on Day 3. Areas of endothelial proliferation and early granulation tissue formation.
On Day 10 of the experiment, histological examination demonstrated pronounced proliferative activity of fibroblast-lineage cells, including tubular proliferation of fibroblasts and vascular endothelial cells. Intact peripheral nerve bundles were identified within the subcutaneous tissue, corresponding to a histological picture of wound epithelialization with preservation of all functional tissue structures (Figure 63).
Figure 63. Experimental Group I. Day 10 of the experiment. Mayer’s hematoxylin and eosin staining. Magnification ×20.
In animals of Group II (vehicle control), edema, lymphohistiocytic infiltration, and degenerative-dystrophic alterations of the subcutaneous tissue and muscle layer persisted through Day 10 of the experiment (Figure 64).
Figure 64. Group II (vehicle control). Day 10 of the experiment. Mayer’s hematoxylin and eosin staining. Magnification ×20.
In Group III (conventional treatment), degenerative-dystrophic changes of the subcutaneous tissue and muscle, together with degenerative alterations of peripheral nerve bundles, remained evident on Day 25 (Figure 65).
Figure 65. Group III (conventional treatment). Day 25 of the experiment. Degenerative changes of nerve bundles. Mayer’s hematoxylin and eosin staining. Magnification ×40.
Histological monitoring demonstrated that granulation tissue formation progressed from the wound margins toward the center of the defect. Consequently, granulation tissue exhibited a focal distribution and considerable cellular polymorphism, consisting of neutrophils, macrophages, and histiocytes. The structural framework of granulation tissue was formed by isolated vertically oriented capillaries of varying diameters, lined by a single layer of endothelial cells. Individual fibroblasts were observed, particularly in deeper wound layers, where they adopted an orientation perpendicular to the newly formed vessels. Poorly differentiated fibroblasts displayed the characteristic spindle-shaped morphology with large nuclei and scant cytoplasm. These cells were distributed throughout the granulation tissue without forming clusters. The tissue itself appeared edematous, with prominent leukocytic infiltration extending from superficial layers to the wound base. Simultaneously with granulation tissue formation, epithelialization was initiated. The degree of epithelial regeneration was closely associated with the extent of granulation. Epithelial repair proceeded through three sequential processes: cell migration, proliferation, and differentiation. The advancing epithelial tongue assumed a wedge-shaped configuration, tapering toward the wound center and exhibiting only mild hypertrophy at the interface with intact skin. The focal distribution of newly formed tissue and its limited surface area corresponded to a relatively short extent of epithelial regeneration.
In animals of Experimental Group I, post-traumatic repair followed a substantially more favorable course, reflected in the morphology of all components of the full-thickness wound defect. The scab was heterogeneous, composed primarily of cellular detritus without plasma-filled regions. The leukocytic barrier consisted predominantly of neutrophils undergoing physiological degeneration. Granulation tissue remained focal in distribution but occupied a considerably larger area than in the control groups. Newly formed tissue was characterized by extensive vascular development and marked fibroblast proliferation. Fibroblastic cells appeared as large, elongated, multiprocess cells, indicating high functional activity. These cells were oriented parallel to the wound surface, particularly in peripheral regions and deeper layers of the wound defect. The extracellular matrix demonstrated progressive accumulation of collagen fibers, which aggregated into bundles.
Thus, treatment with the xenogeneic active substance (Group I) resulted in uniform connective tissue formation throughout the wound defect, distinguishing it from all control groups. Epithelialization proceeded successfully, with complete epidermal differentiation and no evidence of dystrophic alterations. The subcutaneous adipose tissue was not involved in the inflammatory process. Dermal restructuring occurred through progressive granulation tissue formation and stimulation of migration and proliferation of fibroblasts, keratinocytes, endothelial cells, and other reparative cell populations. These processes facilitated tissue restoration, shortened healing time, and prevented complications.
Immunohistochemical analysis was performed on paraffin sections using appropriate primary antibodies and the EnVision visualization system. Cell nuclei were counterstained with hematoxylin. Antigen retrieval was achieved by microwave heat treatment. Microscopic evaluation was conducted using an Olympus CX41 microscope equipped with a CDx41 digital camera and integrated imaging software.
The proliferative activity of epidermal and dermal cells was assessed using monoclonal antibodies against Ki67 (MIB-1), which identify a nuclear antigen present in the majority of proliferating cells. Ki67 is a short-lived protein that undergoes degradation within approximately 1–1.5 hours and is therefore detected exclusively in actively dividing cells. Apoptotic processes were evaluated using monoclonal antibodies directed against Fas receptors (CD95/Apo-1).
During wound healing, particularly throughout the purulent-necrotic phase, the predominant cellular population consisted of neutrophils, ensuring active interaction between the host immune system and bacterial flora. To further characterize immune activity, cells expressing the CD4+ receptor were identified. The overall number of CD4+ cells was reduced compared with intact skin (9.33 ± 0.26 vs. 13.85 ± 0.19, respectively).
CD4 receptors are primarily expressed on T lymphocytes and facilitate interaction with HLA class II antigens, thereby mediating communication between T cells and the macrophage-histiocytic microenvironment. During wound healing, a relative depletion of perivascular T cells was observed compared with normal skin.
A reduction in the number of CD68+ Langerhans cells was also observed (25.7 ± 0.29 vs. 34.0 ± 0.32 in normal skin), together with a diminished capacity for intercellular communication during adaptive immune responses. Migration of CD68+ cells was reduced, accompanied by decreased expression of adhesion molecules, major histocompatibility complex class II products, and costimulatory molecules.
Disruption of physiological regeneration and the appearance of hypertrophic areas appeared to be associated with impaired antigen-presenting function of Langerhans cells and their displacement into the connective tissue underlying the epithelium. These changes suggest defective antigen presentation within skin structures, followed by diminished regulation of physiological and reparative regeneration and initiation of adaptive hypertrophic responses aimed at preserving epithelial barrier function. Concurrently, a reduction in the mast-cell population (CD204+) was observed (2.17 ± 0.2 vs. 4.0 ± 0.18 in controls), accompanied by degranulation. The number of CD14+ skin monocytes increased significantly (11.5 ± 1.4 in wounds vs. 4.34 ± 0.49 in controls).
Within the wound area and adjacent connective tissue, activated CD163+ macrophages and fibroblasts responsible for tissue remodeling were identified. To maximize protection of the wound surface and support complete regeneration, CD8+ lymphocytes emerged, producing cytokines similar to those secreted by CD4+ Th1 cells (TNF-α, TNF-β, IL-2, IL-3, IFN-γ, and GM-CSF). These findings emphasize the crucial role of skin-resident immune and stromal cells during reparative regeneration of infected wounds. Specifically, suppression of the T-cell compartment together with reduced numbers of Langerhans cells and mast cells during the inflammatory phase resulted in impaired antigen presentation, disruption of immune-cell interactions, and subsequent disturbances in normal epidermal and dermal regeneration.
Furthermore, in Experimental Group I, administration of the active xenogeneic substance induced an increase in CD95+ cell carriers during Days 1–3, followed by a gradual decline beginning on Days 3–5. Simultaneously, expression of the proliferation marker Ki67+ increased from the first day onward and remained elevated throughout the 15-day observation period. Between Days 3 and 5, a distinct “marker crossover” occurred, indicating the transition from the inflammatory phase of wound healing to the proliferative phase—that is, completion of cell-death processes and initiation of active tissue regeneration. In contrast, conventionally treated animals demonstrated progressive reduction of apoptosis-related markers, while proliferation activity peaked briefly around Day 5 before declining almost to baseline. Within scar tissue epidermis, active proliferative processes persisted. Ki67 labeling was localized predominantly within nuclei of basal and spinous-layer keratinocytes. In the dermis, positive staining was detected in rounded cells located within blood vessels and between collagen fibers of the regenerative zone, likely representing fibroblast-lineage cells. Expression of Fas protein, indicating readiness of epidermal cells to undergo apoptosis, was identified in basal and spinous-layer keratinocytes, whereas no dermal expression was detected.
The fivefold acceleration of wound cleansing during the inflammatory phase observed in Experimental Group I, together with scar-free epithelialization and restoration of tissue function (as evidenced by hair regrowth), appears to be associated with the ability of the xenogeneic active substance to modulate cellular death pathways during the initial stage of wound healing and to potentiate mechanisms of reparative regeneration. These observations were further supported by immunohistochemical findings. Prior to treatment, expression levels of CD95+ and Ki67+ cells were comparable across all experimental groups. The dynamics of CD95+ and Ki67+ expression are summarized in Tables 9 and 10.
Table 9. Dynamics of the Pro-apoptotic Marker CD95+
Before treatment, all groups exhibited similar baseline CD95+ expression. On Day 1, animals in Group I demonstrated a twofold increase in CD95+ expression (10%), which remained elevated through Day 5 (5–8%).
Table 10. Dynamics of the Proliferation Marker Ki67+
Beginning on Day 3, Experimental Group I exhibited progressive elevation of Ki67+ expression (4–5%), which remained high through Day 15 (8–11%). In Group II (vehicle control), only modest increases in CD95+ and Ki67+ were observed. In Group III (conventional treatment), marker levels remained near baseline during Days 3–5. By Day 15, Ki67+ expression remained high in Experimental Group I, whereas CD95+ and Ki67+ expression in all control groups approached zero (0–4%).
Assessment of activated CD95+ lymphocytes revealed comparable baseline levels across all groups prior to treatment (Figure 66). Likewise, baseline expression of the proliferation marker Ki67+ was similar in all groups before treatment initiation (Figure 67).
In Experimental Group I, treatment induced a transient increase in CD95+ carriers during Days 1–3, followed by gradual decline beginning on Days 3–5. Simultaneously, Ki67+ expression increased from the first day onward and remained elevated throughout the study. The crossover of CD95+ and Ki67+ expression curves between Days 3 and 5 (Figure 68) marked the transition from the inflammatory to the proliferative phase of wound healing.
In Group II (vehicle control), both apoptosis and proliferation markers declined sharply by Days 3–5 (Figure 69). In Group III (conventional treatment), CD95+ expression progressively decreased, while Ki67+ expression exhibited only a transient increase around Day 5 followed by decline toward baseline (Figure 70). Similar suppression of both markers was observed in Group IV (untreated controls) by Days 3–5 (Figure 71).
These findings indicate that the investigated substance acts as an inducer of apoptosis during the inflammatory phase of wound healing and subsequently promotes reparative and proliferative mechanisms during the proliferative and remodeling phases.
Additional experiments demonstrated that a measurable therapeutic effect required the cellular suspension—the source of adhesion molecules—to contain a defined concentration of CD34+CD45dim cells. Reduction of this population below 0.75 × 10⁶ cells/mL significantly diminished reparative efficacy, whereas increasing the concentration above 1.25 × 10⁶ cells/mL produced no meaningful additional benefit. These findings suggest that modulation of apoptosis and proliferation through this cellular phenotype may provide new opportunities for regulation of wound healing, including treatment of infected wounds.
Before proceeding to clinical studies, comprehensive preclinical evaluation of the finished biotechnological product was undertaken. The formulation consisted of SPF chicken embryo cell supernatant collected on Day 6 of gestation, corresponding to a concentration equivalent to 0.75 × 10⁶–1.25 × 10⁶ CD34+CD45dim cells per milliliter, incorporated into a hydroxyethylcellulose-based gel polymer.
Assessment of toxicological safety, microbial contamination, skin-resorptive effects, and antibacterial activity was performed independently in two accredited laboratories. Analytical testing included atomic absorption spectrometry using PinAAcle 900F and MGA-915M systems equipped with the RGT-915 mercury-hydride attachment, as well as gas chromatography performed on GC 2010 Plus and Kristall-2000M instruments. Results were documented in Laboratory Test Report No. 374 KH issued by the Federal Agency for Technical Regulation and Metrology (June 21, 2021) and Laboratory Test Report No. 62L/Z-31.01/23 issued by the LIGHT GROUP testing laboratory (January 31, 2023).
Antibacterial activity was additionally assessed using the standard agar diffusion disk method (Figure 72).
The biotechnological preparation demonstrated significantly larger inhibition zones against Staphylococcus aureus 155, Escherichia coli 2290, and Bacillus cereus JP 5832 than comparator broad-spectrum antibiotics (Tables 11–13).
Thus, the biotechnological agent produced by the author's method functions as a catalyst of granulation tissue growth. It was shown to be safe, sterile, non-toxic, capable of initiating proliferative processes, and additionally possesses bacteriostatic activity.
Considering that wound healing is governed by the same fundamental mechanisms of proliferation and apoptosis and regulated through cell–cell and cell–matrix signaling pathways, together with evidence of dose-dependent effects on cellular viability and apoptosis in culture systems, a biotechnological wound-healing product—Cellgel—was subsequently developed. Its mechanism is based on activation of skin cells exhibiting the CD34+CD45dim phenotype through selective modulation by adhesion molecules.
The results of these independent investigations formed the basis for a series of subsequent scientific studies and for practical application of the identified biological properties. Contract manufacturing of the developed product was organized, and its use was supported by Customs Union Declarations of Conformity No. ТС RU Д-RU.АЛ14.В.03012 (December 25, 2012) and No. ЕАЭС RU Д-RU.РА01.В.56185/21 (June 25, 2021). This enabled implementation of the biotechnological product in dermatological practice for enhancement of reparative processes in erosive and ulcerative skin lesions associated with immune-mediated dermatoses and skin injuries of various etiologies.
Patient I., 52 years old, was admitted to the inpatient department of the Tyumen Regional Dermatovenerologic Dispensary with complaints of intense pruritus, erythema, marked skin dryness, excoriations, edema in the flexural areas of major joints accompanied by fissuring, and extensive scaling of the trunk skin.
According to the medical history, the first manifestations of skin disease appeared in childhood, when a diagnosis of neurodermatitis was established. The disease followed a relapsing course with three or more exacerbations annually, predominantly during the spring and summer months. Subsequently, the diagnosis was revised to atopic dermatitis. Treatment included topical glucocorticosteroids, calcineurin inhibitors, antihistamines, moisturizers, and emollients. During the preceding three years, the disease relapsed more than four times annually and was accompanied by severe pruritus and secondary infection of excoriations. The patient had repeatedly undergone inpatient treatment, including intravenous prednisolone up to 60 mg and antibacterial therapy for infectious complications. The therapeutic effect was considered satisfactory.
Upon admission, the patient's condition was of moderate severity. He had a hypersthenic body habitus. Allergological and hereditary histories were unremarkable. The abdomen was soft and non-tender on palpation. The liver and spleen were not enlarged. Stool was regular and without pathological admixtures. Costovertebral angle tenderness was absent bilaterally. Urination was painless. The thyroid gland was not visibly enlarged and was non-tender on palpation. Lymph nodes were palpable along the posterior cervical chain and in the axillary regions; they were painless, mobile, and soft-elastic in consistency. Nasal breathing was unobstructed. Percussion of the lungs revealed clear pulmonary resonance throughout. Vesicular breath sounds were present without wheezes. Respiratory rate was 18 breaths per minute. Heart sounds were clear and rhythmic, heart rate 74 beats per minute, blood pressure 130/80 mmHg. The tongue was moist and pink. Electrocardiography demonstrated sinus rhythm with a heart rate of 78 beats per minute. Computed tomography revealed no focal or infiltrative pulmonary abnormalities. Ultrasonography of the abdominal organs, retroperitoneal space, thyroid gland, and parathyroid glands showed no pathological findings.
The pathological process was widespread. It was characterized by diffuse erythema of the trunk and extremities, fissures in the antecubital and popliteal fossae, extensive lichenoid scaling with linear excoriations over the entire skin surface, without signs of pyoderma. Cheilitis was present along the vermilion border of the lips. The fingernails exhibited a polished appearance. The SCORAD index was 84.
Laboratory and instrumental investigations on admission revealed normal complete blood count and urinalysis results. Biochemical blood parameters were within reference ranges. Thyroid hormone levels were normal. Serum immunoglobulins A, M, and G were within normal limits. C-reactive protein was 8 mg/L. Total IgE exceeded 1000 IU/mL. Screening for helminthic infestation was negative.
Based on the patient's complaints, disease history, clinical findings, and examination results, the diagnosis was established as:
Atopic dermatitis, generalized form, severe course, active phase with pronounced clinical manifestations, complicated by traumatic excoriations.
During hospitalization, treatment included methylprednisolone 250 mg intravenously in 200 mL of saline solution for seven days, loratadine 10 mg orally for seven days, and topical mometasone therapy. The therapeutic response was limited: pruritus decreased and skin infiltration in the antecubital and popliteal regions diminished; however, numerous excoriations and exudative fissures persisted. This served as the rationale for precision diagnostic evaluation using a skin cytoimmunogram (Table 14).
| Phenotype | Percentage (%) |
|---|---|
| Keratinocytes (CD49f+) | 57.2 |
| Activated keratinocytes (CD49f+ HLA-DR+) | 31.6 |
| Fibroblasts (CD45− CD14− CD44+) | 52 |
| Activated fibroblasts (CD45− CD14− CD44+ CD80+) | 6.7 |
| Langerhans cells (CD207+) | 49 |
| CD207+ CD80− HLA-DR+ | 19.9 |
| CD207+ CD80+ HLA-DR− | 26.0 |
| CD207+ CD80+ HLA-DR+ | 3.1 |
| Endothelial cells (CD146+) | 13.9 |
| CD146+ CD54− HLA-DR+ | 4.2 |
| CD146+ CD54+ HLA-DR− | 9.5 |
| CD146+ CD54+ HLA-DR+ | 0 |
| CD146+ CD34+ | 0.2 |
| Mast cells (CD249+) | 6.6 |
| Activated mast cells (CD249+ CD63+) | 2.9 |
| Monocytes (CD45+ CD14+) | 3.0 |
| Activated monocytes (CD45+ CD14+ HLA-DR+) | 1.5 |
| Total T cells (CD45+ CD3+) | 25 |
| T helper cells (CD4+) | 12 |
| Cytotoxic T cells (CD8+) | 21 |
| B lymphocytes (CD19+) | 6 |
| NK cells (CD16+ CD56+) | 10 |
| Cell viability | 94 |
Overall cell viability was 94%, confirming appropriate sample preparation and reliable phenotypic analysis. The immune phenotype demonstrated a highly active inflammatory pattern with a Th2-dominant profile, pronounced antigen presentation, and involvement of both epidermal and endothelial inflammatory components. The phenotype corresponded to the acute-to-subacute exacerbation phase of atopic dermatitis, characterized by active cellular immune responses and disruption of skin barrier function.
Keratinocytes (CD49f+ HLA-DR+).
The total keratinocyte population constituted 57.2%, with 31.6% exhibiting activation. This marked epidermal activation reflects barrier disruption and enhanced HLA-DR expression. Such hyperactivation is characteristic of atopic dermatitis, in which keratinocytes become not only structural but also immunocompetent cells, producing IL-1, IL-33, and TSLP and initiating Th2-mediated inflammation.
Fibroblasts (CD45− CD14− CD44+ CD80+).
The total fibroblast population was 52%, with 6.7% activated. This moderately elevated fibroblast activity reflects dermal remodeling and participation in inflammatory-reparative processes. Activation is associated with increased production of IL-6 and IL-8 and contributes to the maintenance of inflammatory infiltrates.
Langerhans Cells (CD207+).
The total proportion was 49%, while activated phenotypes (CD80+ HLA-DR+/−) accounted for approximately 50% of the population. This pronounced antigen-presenting activity indicates active involvement of dendritic cells in initiating Th2 responses and is characteristic of acute atopic inflammation.
Endothelial Cells (CD146+ CD54+ CD34+).
The total endothelial population constituted 13.9%, predominantly represented by CD54+ HLA-DR− cells (9.5%). Enhanced endothelial activation, reflected by increased ICAM-1 expression, facilitates leukocyte migration into inflammatory lesions and characterizes vascular hyperreactivity and microcirculatory dysregulation.
Mast Cells (CD249+ CD63+).
The total mast cell population was 6.6%, with 2.9% activated. These cells exhibited moderate degranulation with release of histamine and tryptase, contributing to pruritus and vascular reactivity and amplifying Th2-dominant inflammation.
Monocytes (CD45+ CD14+ HLA-DR+).
Monocytes accounted for 3%, with 1.5% activated. This finding reflects moderate involvement of the phagocytic compartment and ongoing maintenance of inflammatory activity without effective resolution.
Epidermal Lymphocytes
Total T cells (CD3+): 25%
T helper cells (CD4+): 12%
Cytotoxic T cells (CD8+): 21%
B lymphocytes (CD19+): 6%
NK cells (CD16+ CD56+): 10%
The lymphocyte profile demonstrated a moderate Th2/Th1 balance with predominance of cytotoxic T cells and elevated NK-cell activity, consistent with atopic dermatitis involving both adaptive and innate immune responses secondary to epidermal barrier disruption.
The patient's skin phenotype represented an acute inflammatory state characterized by strong antigen presentation, endothelial activation, and partial reparative activity.
Key phenotypic markers included:
significant keratinocyte activation (HLA-DR+) reflecting barrier dysfunction;
hyperactivation of Langerhans cells with increased CD80 expression and initiation of Th2 responses;
endothelial activation (increased ICAM-1 expression), supporting inflammatory cell recruitment;
moderate mast-cell degranulation;
preserved fibroblast activity contributing to tissue remodeling;
Th2-skewed immune imbalance amplified by innate immune effectors (NK cells).
This cytoimmunological profile corresponded to a decompensated inflammatory phenotype.
Skin Cytoimmunogram of Patient I. Before Treatment. The skin cytoimmunogram obtained before treatment demonstrated a highly active inflammatory state characterized by a Th2-dominant immune profile and disruption of the epidermal barrier.
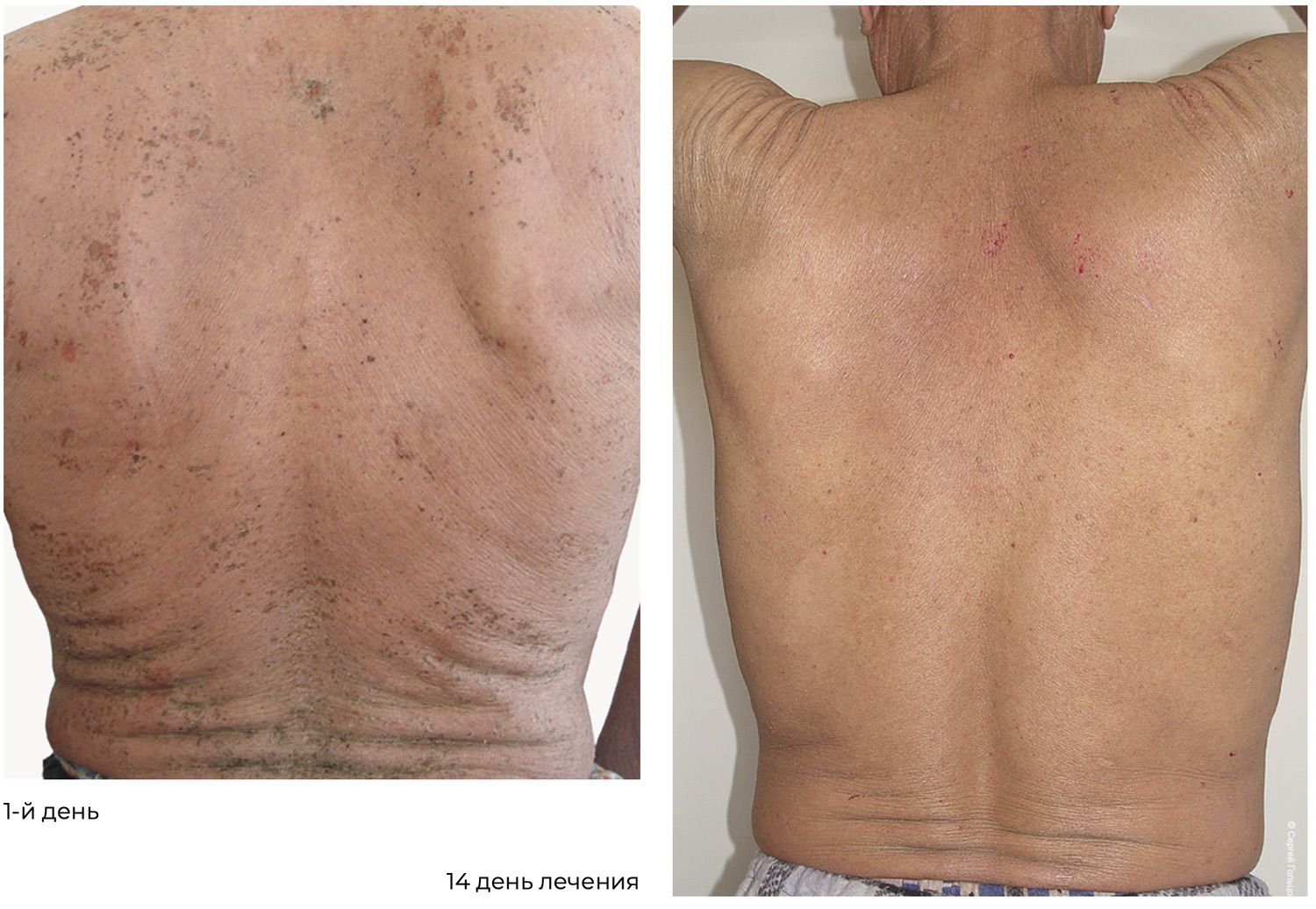
The principal pathogenic mechanisms included:
hyperactivation of the epidermis and dendritic cells;
vascular dysregulation and leukocytic infiltration;
preserved reparative activity of fibroblasts;
mast-cell and NK-cell participation in inflammation.
This phenotype corresponded to an acute inflammatory type and represented a diagnostically meaningful reference point for monitoring therapeutic efficacy through comparison with subsequent skin cytoimmunograms.
Because the cytoimmunogram revealed the presence of endothelial cells with preserved reparative potential but insufficient regenerative activation, as reflected by the CD146+ and CD146+CD34+ populations, Cellgel biotechnological gel was added to topical mometasone therapy. The preparation was applied twice daily for 10 days to erosive lesions and skin fissures to prevent secondary infection and accelerate epithelialization.
A favorable clinical response was observed. Along with a reduction in the total area of skin involvement and a marked decrease in lichenification, secondary lesions—including erosions and crusted scales—regressed significantly. No adverse events were recorded. According to the patient, he had not experienced such a degree of “skin comfort” for more than ten years.
Simultaneously, the clinical improvement, reflected by a decrease in the SCORAD index from 84 to 22 points, was accompanied by a reduction in inflammatory cell populations and a marked increase in activated endothelial cells involved in tissue repair. Specifically, the proportion of CD146+CD34+ endothelial cells increased from 0.2% before treatment to 8.9% after treatment (Table 15).
| Phenotype | Percentage (%) |
|---|---|
| Keratinocytes (CD49f+) | 54.1 |
| Activated keratinocytes (CD49f+ HLA-DR+) | 24.3 |
| Fibroblasts (CD45− CD14− CD44+) | 48 |
| Activated fibroblasts (CD45− CD14− CD44+ CD80+) | 2.7 |
| Langerhans cells (CD207+) | 39 |
| CD207+ CD80− HLA-DR+ | 14.3 |
| CD207+ CD80+ HLA-DR− | 16.0 |
| CD207+ CD80+ HLA-DR+ | 8.7 |
| Endothelial cells (CD146+) | 25.4 |
| CD146+ CD54− HLA-DR+ | 5.2 |
| CD146+ CD54+ HLA-DR− | 11.3 |
| CD146+ CD54+ HLA-DR+ | 0 |
| CD146+ CD34+ | 8.9 |
| Mast cells (CD249+) | 2.6 |
| Activated mast cells (CD249+ CD63+) | 0.9 |
| Monocytes (CD45+ CD14+) | 2.0 |
| Activated monocytes (CD45+ CD14+ HLA-DR+) | 0.5 |
| Total T cells (CD45+ CD3+) | 15 |
| T-helper cells (CD4+) | 9 |
| Cytotoxic T cells (CD8+) | 11 |
| B lymphocytes (CD19+) | 8 |
| NK cells (CD16+ CD56+) | 8 |
| Cell viability | 95 |
Overall cell viability increased to 95%, indicating restoration of tissue homeostasis. The immune profile demonstrated attenuation of the inflammatory phenotype and a transition from an acute Th2-dominant inflammatory state toward a balanced reparative-homeostatic phenotype.
Compared with the baseline cytoimmunogram, there was a reduction in antigen-presenting cell activation, normalization of the vascular component, and restoration of fibroblast activity, all of which suggested recovery of both barrier and structural skin functions.
The total keratinocyte population was 54.1%, with 24.3% activated. Compared with baseline values (31.6%), activated keratinocytes decreased substantially, reflecting restoration of barrier function and reduced cytokine-induced HLA-DR expression. The epidermis retained regenerative potential without evidence of hyperreactivity.
Fibroblasts constituted 48% of the cell population, with 2.7% activated. Although the total fibroblast population decreased slightly from 52%, activated fibroblasts declined by more than half (from 6.7% to 2.7%), indicating completion of the active reparative phase and stabilization of the dermal extracellular matrix.
The overall proportion of Langerhans cells decreased to 39%, while activated phenotypes (CD80+ HLA-DR+/−) declined markedly compared with baseline values. This reduction reflected attenuation of antigen-presenting activity and decreased migration of dendritic cells into the epidermis.
The total endothelial cell population increased to 25.4%, with a pronounced shift toward the CD34+ subset (8.9%), a marker of neovascularization and microvascular repair. Activated endothelial phenotypes declined, indicating a transition from inflammatory vascular activation toward regenerative angiogenesis.
Mast cells accounted for 2.6% of the population, with only 0.9% activated. Degranulation activity decreased to approximately one-third of baseline levels, corresponding clinically to diminished pruritus and vascular hyperreactivity.
Activated monocytes decreased from 1.5% to 0.5%, reflecting resolution of inflammatory activity and normalization of phagocytic function.
Total T cells (CD3+): 15%
T-helper cells (CD4+): 9%
Cytotoxic T cells (CD8+): 11%
B lymphocytes (CD19+): 8%
NK cells (CD16+ CD56+): 8%
The CD4/CD8 ratio normalized from approximately 0.6 before treatment to 0.8 after treatment, indicating restoration of Th1/Th2 balance and resolution of Th2-dominant inflammation. B-cell and NK-cell populations remained stable, consistent with physiologic innate immune activity.
Following therapy, the cytoimmunogram demonstrated restoration of cellular homeostasis and substantial reduction of inflammatory activity. The principal phenotypic features included:
decreased activation of antigen-presenting cells;
normalization of endothelial responses;
restoration of epidermal barrier integrity;
reduced mast-cell degranulation;
stabilization of the Th1/Th2 immune balance.
The skin phenotype could therefore be classified as a post-inflammatory reparative phenotype, corresponding to clinical remission of atopic dermatitis.
From the perspective of Phenotypic Dermatology, this case illustrates a transition from a dysregulated Th2-inflammatory phenotype characterized by hyperactivation of antigen-presenting cells, endothelial dysfunction, and mast-cell degranulation, to a structurally balanced phenotype focused on maintenance of barrier integrity and angiogenic repair. The skin thus shifted from a state of chronic immune activation toward functional stability and restoration of homeostatic balance.
Importantly, these findings suggest a close immunopathogenetic relationship between the biological components of Cellgel and the cellular characteristics identified through precision analysis of inflammatory infiltrates in atopic dermatitis. This relationship appears to underlie the observed reparative effects and may explain the clinical efficacy of Cellgel as an adjunct to topical therapy. Furthermore, the positive outcome supports future investigation of Cellgel in patients with severe atopic dermatitis complicated by erosions and excoriations.
To optimize topical therapy, an experimental study was conducted to determine whether the developed biotechnological product possessed reparative activity in erosive and ulcerative skin lesions of patients with pemphigus vulgaris. This work resulted in Russian Federation Patent No. 2481115, granted on March 23, 2012.
Patient Ya., a 55-year-old woman, presented to the Tyumen Regional Dermatovenerologic Dispensary in December 2018 with complaints of widespread symmetrical eruptions on the skin of the trunk and extremities. According to the patient, the disease had begun acutely during the summer of that year. After the appearance of the first signs—isolated erythematous lesions on the trunk accompanied by moderate pruritus—she consulted a dermatologist at her local clinic. A provisional diagnosis of “Dermatitis?” was established, and symptomatic therapy with a combined glucocorticosteroid preparation was prescribed. Several days later, vesicular lesions began to appear within the erythematous areas. She was subsequently hospitalized for diagnostic clarification. Her medical history, family history, and allergy history were unremarkable.
At admission, the pathological process was widespread, involving the trunk and both upper and lower extremities. The lesions consisted of multiple round, partially edematous erythematous-squamous plaques of pink and pale-pink coloration, accompanied by areas of brown hyperpigmentation. On the surface of certain lesions, particularly on the chest, back, and lumbar region, grouped vesicles containing serous fluid were observed on a hyperemic background. On the lateral surfaces of the trunk, buttocks, and upper third of the arms, coalescing oval and round erosions up to 1 cm in diameter were present, covered with crusts and scales. Nikolsky’s sign was positive. Skin appendages and mucous membranes were not involved.
The preliminary clinical diagnosis included:
Pemphigus vulgaris?
Pemphigus foliaceus?
Dermatitis herpetiformis (Duhring disease)?
Given the patient’s severe condition, the rapid development of vesicular and erosive lesions, her age, and the positive Nikolsky sign, a diagnostic skin biopsy was obtained from the back for histopathological examination. Histological findings necessitated additional diagnostic investigations. Specifically:
“The epidermis was irregular, with pronounced hyperkeratosis and spongiosis; cells of the granular layer were separated from one another with a tendency toward blister formation. The papillary dermis exhibited edema and diffuse lymphocytic infiltration with eosinophils.”
Based on these findings, direct immunofluorescence studies and a skin cytoimmunogram were recommended.
Comprehensive clinical and laboratory evaluation revealed:
a positive Tzanck smear demonstrating acantholytic cells;
peripheral blood leukocytosis (12.8 × 10⁹/L);
lymphopenia (1.9 thousand/μL);
elevated ALT (72 U/L);
indirect immunofluorescence showing moderate IgG deposition within intercellular spaces of the epidermis, with decreasing intensity from the granular and cornified layers toward the basal layer.
Additional information obtained through analysis of the skin cytoimmunogram demonstrated the presence of reparative potential within inflammatory lesions (Table 16).
| Phenotype | Percentage (%) |
|---|---|
| Keratinocytes (CD49f+) | 41.2 |
| Activated keratinocytes (CD49f+ HLA-DR+) | 26.2 |
| Fibroblasts (CD45− CD14− CD44+) | 31.0 |
| Activated fibroblasts (CD45− CD14− CD44+ CD80+) | 2.9 |
| Langerhans cells (CD207+) | 54 |
| CD207+ CD80− HLA-DR+ | 22.0 |
| CD207+ CD80+ HLA-DR− | 19.0 |
| CD207+ CD80+ HLA-DR+ | 3.0 |
| Endothelial cells (CD146+) | 15.1 |
| CD146+ CD54− HLA-DR+ | 3.2 |
| CD146+ CD54+ HLA-DR− | 7.5 |
| CD146+ CD54+ HLA-DR+ | 3.3 |
| CD146+ CD34+ | 1.1 |
| Mast cells (CD249+) | 13.1 |
| Activated mast cells (CD249+ CD63+) | 1.9 |
| Monocytes (CD45+ CD14+) | 2.0 |
| Activated monocytes (CD45+ CD14+ HLA-DR+) | 0.5 |
| Total T cells (CD45+ CD3+) | 29 |
| T-helper cells (CD4+) | 11 |
| Cytotoxic T cells (CD8+) | 17 |
| B lymphocytes (CD19+) | 0 |
| NK cells (CD16+ CD56+) | 1 |
| Cell viability | 96 |
Overall cell viability was 96%, indicating excellent preservation of biopsy material despite severe inflammatory and destructive skin changes. The profile reflected a pronounced autoimmune inflammatory process with activation of antigen-presenting and epidermal cells characteristic of pemphigus. The immune phenotype demonstrated:
keratinocyte hyperactivation;
activation of dendritic and endothelial cells;
extensive antigen presentation;
enhancement of the cytotoxic immune compartment.
This phenotype corresponded to a decompensated autoimmune inflammatory state, characterized by imbalance between antigen presentation, tissue destruction, and impaired barrier function.
Keratinocytes constituted 41.2% of the cell population, with 26.2% expressing HLA-DR. Elevated HLA-DR expression reflected active participation of keratinocytes in antigen presentation and autoimmune amplification. Such a profile is typical of pemphigus vulgaris, in which keratinocyte activation is associated with autoantigen production (desmogleins 1 and 3), leading to loss of desmosomal adhesion and intraepidermal blister formation.
Fibroblasts accounted for 31% of cells, with 2.9% activated. Compared with keratinocytes, connective-tissue activation was less pronounced, reflecting a secondary dermal response to tissue injury. Activated fibroblasts likely contributed to production of TGF-β and IL-6, supporting angiogenesis and inflammatory signaling.
Langerhans cells comprised 54% of the total population, with activated phenotypes representing approximately 45%. This indicated excessive antigen presentation and migration of dendritic cells toward regions of autoimmune injury. Such activation is characteristic of pemphigus, where dendritic cells interact closely with T lymphocytes and promote autoantibody production.
Endothelial activation was evident, supporting increased vascular permeability and exudation into blister cavities. The presence of CD34+ cells suggested compensatory angiogenic activity in response to inflammatory tissue destruction.
Mast cells represented 13.1% of the population, while activated forms accounted for 1.9%, indicating mild degranulation typical of chronic autoimmune inflammation.
Monocytes were present at low levels (2%), likely reflecting migration into lesions and differentiation into tissue macrophages.
The lymphocyte profile included:
T-helper cells (CD4+): 11%
Cytotoxic T cells (CD8+): 17%
B lymphocytes (CD19+): 0%
NK cells: 1%
The predominance of CD8+ cells indicated a significant cytotoxic component of autoimmune skin injury, whereas the absence of B cells in the biopsy was consistent with their primary localization in the circulation.
Overall, the cytoimmunogram reflected an active autoimmune inflammatory phase of pemphigus vulgaris, dominated by activation of keratinocytes, Langerhans cells, and cytotoxic T cells. The phenotype corresponded to uncontrolled inflammation with destruction of tissue architecture and warranted phenotypic monitoring during immunosuppressive treatment.
The patient received the following inpatient treatment:
oral prednisolone at 1 mg/kg/day (90 mg/day) for 30 days;
intravenous essential phospholipids, 5 mL, five administrations;
Panangin®, one tablet three times daily for 30 days;
Calcium D3®, one tablet three times daily;
topical mometasone furoate;
topical methylene blue solution.
Given the reparative potential identified by the presence of CD146+CD34+ cells (1.1%), Cellgel was additionally recommended for application to erosive lesions twice daily for 30 days to accelerate epithelialization.
During therapy, marked clinical improvement was observed, including regression of most lesions with residual hyperpigmentation. The patient was discharged with improvement and continued outpatient follow-up under the supervision of a local dermatologist.
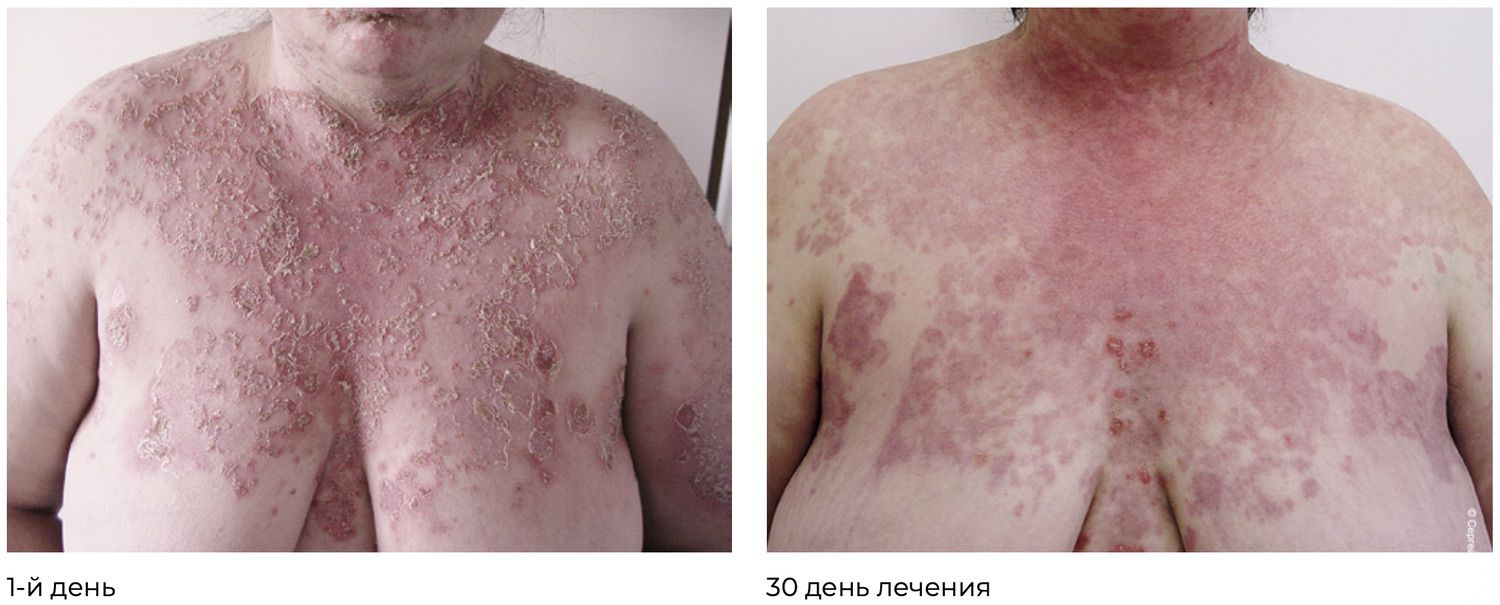
At subsequent follow-up, sustained clinical remission was documented, with no new lesions appearing. Two months after hospitalization, the daily prednisolone dose was reduced from 60 mg/day at discharge to 45 mg/day.
Continuing in the same strict scientific American academic style:
It was established (Table 17) that a close immunopathogenetic relationship exists among the constituent components of the developed biotechnological product, providing a combined synergistic proliferative and anti-apoptotic effect and thereby accounting for its high therapeutic efficacy.
| Phenotype | Percentage (%) |
|---|---|
| Keratinocytes (CD49f+) | 63.2 |
| Activated keratinocytes (CD49f+ HLA-DR+) | 39.2 |
| Fibroblasts (CD45– CD14– CD44+) | 42.0 |
| Activated fibroblasts (CD45– CD14– CD44+ CD80+) | 6.9 |
| Langerhans cells (CD207+) | 29 |
| CD207+ CD80– HLA-DR+ | 18.0 |
| CD207+ CD80+ HLA-DR– | 11.0 |
| CD207+ CD80+ HLA-DR+ | 1.0 |
| Endothelial cells (CD146+) | 21.1 |
| CD146+ CD54– HLA-DR+ | 9.2 |
| CD146+ CD54+ HLA-DR– | 1.5 |
| CD146+ CD54+ HLA-DR+ | 4.3 |
| CD146+ CD34+ | 5.1 |
| Mast cells (CD249+) | 10.2 |
| Activated mast cells (CD249+ CD63+) | 0.9 |
| Monocytes (CD45+ CD14+) | 2.0 |
| Activated monocytes (CD45+ CD14+ HLA-DR+) | 1.5 |
| Total T lymphocytes (CD45+ CD3+) | 34 |
| T-helper lymphocytes (CD4+) | 23 |
| Cytotoxic T lymphocytes (CD8+) | 10 |
| B lymphocytes (CD19+) | 0 |
| NK cells (CD16+ CD56+) | 0 |
| Cell viability | 98 |
The pretreatment cytoimmunogram (No. 111/2) reflected pronounced autoimmune inflammation with decompensation of barrier and regulatory mechanisms, characterized by keratinocyte hyperactivation, excessive antigen presentation, vascular hyperreactivity, and enhancement of the cytotoxic immune compartment. Following treatment (skin cytoimmunogram No. 111/3, February 14, 2018), a stable normalization of the cutaneous immune microenvironment was observed, accompanied by a reduction in inflammatory markers and a transition of the phenotype from an autoaggressive to a reparative-homeostatic state.
Cell viability reached 98%, indicating excellent preservation of the cellular suspension and restoration of skin trophic status. The cytoimmune profile demonstrated a marked reduction in inflammatory activity and a shift toward a reparative phenotype.
Keratinocytes (CD49f+) accounted for 63.2% of the population, of which 39.2% expressed HLA-DR. The increase in the overall keratinocyte population, despite moderate activation, reflects restoration of the structural framework of the epidermis and attenuation of autoimmune injury.
Fibroblasts (CD45– CD14– CD44+) represented 42% of the cell population, with 6.9% expressing the activation marker CD80. Expansion of the fibroblast compartment while maintaining relatively low activation supports ongoing reparative processes within the dermis in the absence of pronounced inflammation.
The total proportion of Langerhans cells (CD207+) was 29%, including:
CD80– HLA-DR+ : 18%
CD80+ HLA-DR– : 11%
CD80+ HLA-DR+ : 1%
The overall reduction in Langerhans cell abundance compared with the pretreatment phase (54% versus 29%) indicates normalization of antigen-presenting activity and resolution of the autoimmune reaction within the epidermis.
Endothelial cells (CD146+) comprised 21.1% of the population, with activated forms (CD54+ HLA-DR+) accounting for 4.3%. These findings indicate restoration of endothelial integrity and reduction of vascular inflammatory changes that had previously accompanied the autoimmune process.
Mast cells (CD249+) represented 10.2% of the population, while activated (CD63+) forms accounted for only 0.9%. This marked reduction in degranulated mast cells reflects suppression of allergy-like inflammatory mechanisms and stabilization of membrane receptor function.
Monocytes (CD45+ CD14+) accounted for 2% of the population, with 1.5% expressing HLA-DR. The modest increase in activated forms compared with baseline likely reflects physiological macrophage activity during tissue remodeling and repair.
Total epidermal lymphocytes (CD45+ CD3+) accounted for 34%, including:
T-helper cells (CD4+): 23%
Cytotoxic T cells (CD8+): 10%
B lymphocytes (CD19+): 0%
NK cells (CD3– CD16+ CD56+): 0%
Normalization of the CD4/CD8 ratio, together with the absence of detectable NK and B lymphocytes, indicates resolution of autoimmune activation and transition of the immune response toward a regulatory state.
Following treatment, a clear normalization of the cutaneous cytoimmune profile was observed:
a reduction in antigen-presenting cell populations;
a decrease in activated endothelial and mast-cell fractions;
restoration of T-lymphocyte balance;
expansion of reparative fibroblast populations.
The overall pattern corresponds to the remission phase of pemphigus vulgaris.
The phenotypic dynamics observed in patient Ya. confirm:
the efficacy of therapy aimed at suppressing antigen presentation and autoantibody-mediated responses;
restoration of epidermal histoarchitecture;
normalization of the dermal microenvironment and microcirculation;
sustained reduction of phenotypic inflammatory activity to remission levels.
Accordingly, phenotypic analysis further supports the diagnostic value of the skin cytoimmunogram as a tool for monitoring the dynamics of autoimmune dermatoses.
Thus, the inclusion of the developed biotechnological product Cellgel in the comprehensive topical treatment regimen of patients with pemphigus vulgaris appears well justified. The identification of the CD34+ CD45dim cellular phenotype provided a methodologically rigorous foundation for the development of the wound-healing product Cellgel, which subsequently became a multiple recipient of awards in the national “100 Best Goods of Russia” competition and the regional “100 Best Goods of the Tyumen Region” competition.
The term “amplification,” introduced by René Descartes in his Meditations on First Philosophy, originally denoted the concept of “enhancement” or “intensification.” In the present work, however, the term is used as the name of a technical invention and is interpreted as an activator of the reparative potentials inherent in skin cells, potentials that can be progressively and functionally expressed under appropriate conditions.
Within one of the conceptual frameworks developed in this study, a species-level concept corresponding to a particular skin-cell phenotype was identified. This served as the basis for formulating the task of designing a technical device capable of selectively activating, in vitro, skin cells exhibiting a specific phenotype. Taking into account the results of the conceptualization process, the problem was formulated as follows.
Among the full diversity of phenotypes represented by species-specific cellular subpopulations, each characterized by a defined set of functions within conceptual scheme D3, identify a phenotype formed by groups of cells of different types that share common attributes and, consequently, common functional properties. Such a phenotype is defined by the following expression:
Т4 = {t ∈ B(Х1) | (x1 ∈ t) & (x2 ∈ t) ⇒ ((∃d1 ∈ D) & (∃d2 ∈ D) & (pr1 d1 = x1) & (pr1 d2 = x2) & (pr2 d1 = pr2 d2))}.
Based on this phenotype, develop a technical device capable of selectively activating the functional properties of the corresponding cells.
It was established (Table 18) that a close immunopathogenetic relationship exists among the constituent components of the developed biotechnological product. This relationship provides a combined, synergistic proliferative and anti-apoptotic effect and accounts for the high therapeutic efficacy of the preparation.
Table 18. Skin Cytoimmunogram of Patient Ya., 55 Years Old, February 14, 2018 (No. 111/3)
|
Субпопуляции клеток кожи |
Фенотип |
Активация ксеногенным активатором |
Активация цитокинами |
||
|
75 мкг/мл активатора |
150 мкг/мл активатора |
IL-1 2мкг/мл |
IL-6 2мкг/мл |
||
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
55,0 9,6 |
79,2 40,5 |
80,9 15,6 |
54,0 13,5 |
|
Фибробласты, |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
81,3 10,5 |
74,3 25,1 |
92,6 7,0 |
93,1 6,9 |
|
Клетки Лангерганса, |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
63,1 0,9 1,2 19,6 |
49,2 0,2 6,4 14,2 |
60,9 1,0 10,0 0,6 |
45,3 0 6,4 0 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
2,7 0 20,9 35,1 70,2 |
2,6 1,0 43,1 25,8 69,0 |
0,7 0,2 70,2 7,0 60,2 |
0,8 0 70,6 5,1 59,2 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
3,2 0,9 |
19,5 18,6 |
10,3 15,6 |
7,1 1,0 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
9,5 3,2 |
20,3 10,6 |
5,8 1,0 |
7,0 2,9 |
|
Эпидермальные лимфоциты Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
12,0 9,0 3,0 15,2 10,6 |
14,1 11,3 1,9 14,7 14,0 |
13,2 10,0 3,0 6,9 10,2 |
12,9 10,3 2,4 15,9 11,1 |
a xenogeneic biotechnological activator at concentrations of 75 μg/mL and 150 μg/mL;
the cytokines IL-1 and IL-6 at 2 μg/mL.
Changes in the relative abundance of cells expressing activation-associated phenotypes (CD44+, CD80+, HLA-DR+, CD54+, CD63+, CD14+, etc.) were assessed, allowing evaluation of shifts in cellular homeostasis from a resting state toward reparative activity.
The results demonstrate fundamentally different patterns of response to cytokine stimulation and to the biotechnological complex, findings of considerable significance for the regulation of cutaneous repair processes.
At 75 μg/mL, moderate activation was observed (55% total keratinocytes, 9.6% HLA-DR+).
At 150 μg/mL, pronounced stimulation occurred (79.2% total, 40.5% HLA-DR+).
IL-1 induced the greatest proliferative response (80.9%), while HLA-DR+ activation remained limited (15.6%).
IL-6, in contrast, reduced the overall keratinocyte pool to 54%, while maintaining 13.5% activated cells.
Thus, the xenogeneic activator induces a barrier-reparative activation phenotype in keratinocytes, combining proliferation with moderate antigen-presenting activity. Cytokines exert more selective effects: IL-1 enhances reparative processes, whereas IL-6 promotes normalization and a return to homeostasis.
75 μg/mL: increased CD44+ (81%) and CD80+ (10.5%) populations.
150 μg/mL: enhancement to 25.1% activated CD80+ fibroblasts.
IL-1 and IL-6 maintained high CD44+ levels (92–93%) while keeping CD80+ expression relatively low (~7%).
These findings reflect a balance between stimulation and modulation. The xenogeneic activator induces an active phase of extracellular matrix remodeling—a form of “reparative burst”—whereas cytokines support gradual differentiation and completion of the regenerative process.
Under the xenogeneic activator, CD207+ cells reached 63% at 75 μg/mL and 49% at 150 μg/mL, with activated CD80+ HLA-DR+ cells reaching 14%.
IL-1 and IL-6 produced more limited activation (10% and 6%, respectively).
These data indicate controlled activation of antigen presentation—an important feature of xenogeneic stimulation that initiates regenerative cascades without triggering overt inflammation.
At 75 μg/mL, substantial increases in CD54+ HLA-DR− cells (20.9%) and CD34+ cells (70%) were observed, consistent with active neoangiogenesis.
At 150 μg/mL, CD34+ levels remained high (69%) while activation markers declined.
IL-1 and IL-6 produced a more moderate angiogenic response (CD34+ ≈ 60%).
Accordingly, the xenogeneic product promotes a vascular-reparative pathway, whereas cytokines contribute to stabilization and morphogenetic maturation of the capillary network.
At 75 μg/mL, only minimal changes were observed (3% total, 0.9% activated).
At 150 μg/mL, marked elevation of CD63+ cells (18.6%) indicated mast-cell degranulation and release of growth mediators.
IL-1 and IL-6 attenuated this effect (~10% and ~1%, respectively).
These findings support the concept that the xenogeneic activator initiates the reparative response through a transient inflammatory phase, whereas cytokines serve as physiological modulators during tissue maturation.
At 150 μg/mL, maximal activation was observed (20% total monocytes, 10% HLA-DR+).
IL-1 and IL-6 induced 5–7% CD14+ cells, with low HLA-DR+ expression (<3%).
This pattern is consistent with induction of an M2-oriented macrophage phenotype, supporting tissue clearance, angiogenesis, and extracellular matrix remodeling.
All stimuli moderately increased total T-cell numbers (13–14%).
IL-1 and IL-6 balanced the Th1/Th2 axis (CD4+ ≈ 10%, CD8+ ≈ 3%).
NK-cell activity increased from approximately 10% to 11%, predominantly in response to cytokines, reflecting participation in tissue repair and antimicrobial defense.
Overall, the cellular immune system maintained a homeostatic orientation without evidence of autoimmune activation.
The xenogeneic biotechnological product induces a coordinated and physiologically balanced activation program closely resembling the natural phases of tissue repair:
Initiation (inflammatory–trophic phase) — transient activation of HLA-DR+ and CD80+ phenotypes.
Regeneration (proliferative–vascular phase) — activation of CD34+, CD44+, and CD49f+ populations.
Maturation (normalization–homeostatic phase) — gradual reduction of cytokine stimulation, expansion of fibroblast populations, and differentiation of keratinocytes.
This activation model demonstrates controlled, non-autoimmune skin regeneration, supporting the potential of the xenogeneic biotechnological complex as a universal regulator of tissue repair and cutaneous homeostasis.
|
Субпопуляции клеток кожи |
Фенотип |
Образец ex tempore |
Образец после криоконсервации |
||||
|
% |
TNFα 2мкг/мл |
ИНФγ 2мкг/мл |
% |
TNFα 2мкг/мл |
ИНФγ 2мкг/мл |
||
|
Кератиноциты, из них активированные |
CD49f+ CD49f+ HLA-DR+ |
54,0 6,9 |
58,2 9,3 |
76,6 8,9 |
54,6 1,3 |
56,0 8,3 |
69,4 8,0 |
|
Фибробласты, |
CD45– CD14– CD44+ CD45– CD14– CD44+ CD80+ |
86 6,2 |
82,1 8,9 |
92,6 8,4 |
66,8 3,5 |
79,4 6,2 |
85,1 7,9 |
|
Клетки Лангерганса, |
CD207+ CD207+ CD80– HLA-DR+ CD207+ CD80+ HLA-DR– CD207+ CD80+ HLA-DR+ |
46,1 0 4,9 0,3 |
30,2 0 5,0 0,3 |
64,9 11,6 4,5 1,9 |
46,4 0 1,3 3,5 |
28,3 0 1,9 3,1 |
62,3 5,0 3,9 1,0 |
|
Эндотелиальные клетки, из них активированные |
CD146+ CD146+ CD54– HLA-DR+ CD146+ CD54+ HLA-DR– CD146+ CD54+ HLA-DR+ CD146+ CD34+ |
0,6 0 60,3 5,4 59,0 |
0,9 0,1 15,9 1,2 35,2 |
1,9 1,5 20,0 2,3 62,0 |
0,6 0 50,0 0 36,0 |
0,8 0 16,1 0,9 29,1 |
1,6 0,5 22,3 1,9 44,0 |
|
Тучные клетки, из них активированные |
CD249+ CD249+ CD63+ |
6,8 1,1 |
6,5 1,3 |
10,7 9,8 |
2,5 1,6 |
4,7 1,9 |
9,2 7,1 |
|
Моноциты, из них активированные |
CD45+ CD14+ CD45+ CD14+ HLA-DR+ |
7,0 2,6 |
22,0 6,8 |
24,5 3,8 |
5,6 0 |
16,2 2,6 |
19,3 3,0 |
|
Эпидермальные лимфоциты Т-общие Т-хелперы Т-цитотоксические В-лимфоциты NK-клетки |
CD45+ CD3+ CD45+ CD3+ CD4+ CD8– CD45+ CD3+ CD4– CD8+ CD45+ CD3+ CD19+ CD45+ CD3– CD16+ CD56+ |
13,0 11,9 1,2 8,1 12,5 |
16,2 14,0 2,2 15,3 23,6 |
25,0 15,0 5,0 9,2 18,2 |
11,2 9,9 1,3 7,0 9,5 |
12,9 10,0 2,9 13,2 20,6 |
19,3 13,5 5,8 8,6 16,2 |
The study summarized in Table 19 demonstrates how the xenogeneic biotechnological product modulates the reactivity of the principal human skin-cell subpopulations under stimulation with the pro-inflammatory cytokines TNF-α and IFN-γ, which model conditions of immune stress and tissue repair.
Comparisons were performed between:
native skin samples (ex tempore), and
cryopreserved samples, modeling long-term storage and transplantation-related reconstruction.
In native samples, TNF-α increased the total keratinocyte population to 58.2%, whereas IFN-γ increased it to 76.6%, accompanied by HLA-DR+ expression of 8.9–9.3%.
In cryopreserved samples, values were slightly lower (54–56% total; 8–8.3% activated), although cellular responsiveness was preserved.
Interpretation: Both cytokines induce keratinocyte activation through antigen-presentation pathways. IFN-γ produces a stronger phenotype of immune awakening, whereas TNF-α predominantly stimulates regenerative proliferation. Even after cryopreservation, skin cells retain plasticity and regenerative capacity, underscoring the resilience of the epidermal reserve.
In native samples, fibroblast activity remained high (86%). TNF-α moderately reduced CD44+ cells to 82%, whereas IFN-γ increased them to 92%, while CD80+ expression remained approximately 8–9%.
In cryopreserved samples, the overall fibroblast pool was partially reduced (66–79%), yet IFN-γ maintained high metabolic activity (up to 85%).
These observations suggest that the xenogeneic biomolecular complex provides fibroblasts with a form of “anabolic memory”—the ability to preserve regenerative potential despite cryogenic stress and cytokine exposure.
In native samples, IFN-γ markedly enhanced activation (CD207+ = 64.9%; CD80+ HLA-DR+ ≈ 11%), whereas TNF-α produced a more modest effect (~30%).
Following cryopreservation, responsiveness was partially reduced (CD207+ = 46–62%), although the trend toward increased antigen presentation under IFN-γ stimulation persisted.
Thus, IFN-γ activates the epidermal immune compartment through a Th1-oriented mechanism, while TNF-α supports structural renewal without excessive inflammatory activation.
In native samples, TNF-α induced CD54+ cells up to 15% and CD34+ cells to approximately 35%, whereas IFN-γ enhanced angiogenic characteristics to 62%.
In cryopreserved samples, responses were attenuated but retained the same pattern: IFN-γ increased the CD34+ population to approximately 44%, indicating preservation of angiogenic capacity.
These findings suggest that the xenogeneic product stabilizes the endothelial reserve even after cryopreservation, an important property for restoration of microcirculation during transplantation and in chronic dermatoses.
In native samples, IFN-γ increased mast-cell abundance to approximately 10%, with CD63+ cells reaching 9.8%, whereas TNF-α induced only moderate degranulation.
Following cryopreservation, overall levels decreased, although responsiveness to IFN-γ was maintained (~9%).
This pattern reflects modulation of the para-inflammatory response: the xenogeneic stimulus does not induce excessive degranulation but instead promotes balanced release of growth factors such as VEGF, FGF, and TGF-β.
In native samples, activated monocytes reached 22% with TNF-α and 24% with IFN-γ, while HLA-DR+ expression remained approximately 4%.
After cryopreservation, values declined moderately (16–19%), while maintaining an M2-oriented differentiation profile.
These results further support the role of the xenogeneic complex in establishing an anti-inflammatory macrophage phenotype that promotes repair without sustaining chronic inflammation.
Total T lymphocytes (CD3+) increased from 13% to 25% under IFN-γ stimulation.
T-helper cells (CD4+) were moderately activated (~14–15%).
Cytotoxic T cells (CD8+) remained low (~2–5%), indicating absence of autoaggressive responses.
B lymphocytes and NK cells displayed adaptive activation, increasing by approximately 1.5–2-fold.
Together with the biomolecular complex, these cytokines restore Th1/Th2 balance and stimulate physiological immune surveillance without damaging the epidermis.
TNF-α, in combination with the xenogeneic biotechnological complex, activates a reparative-structural pathway, enhancing keratinocyte and fibroblast proliferation without pronounced immune activation.
IFN-γ induces regulated antigen presentation and angiogenesis, engaging Langerhans cells and endothelial cells in a phase of restorative inflammation.
Cryopreserved samples retain substantial regenerative potential, demonstrating the phenotypic stability of skin tissues under xenogeneic biostimulation.
Collectively, the data indicate that the xenogeneic product creates an optimal cytokine environment for tissue repair by coordinating activation of epidermal, dermal, and immune compartments.
The activation results obtained in these experiments provided the basis for planning an evaluation of the developed amplifier model in a patient with skin injury.
Case 11. Amplification of the Reparative Potential of Skin in the Treatment of a Grade IIIA Burn
Patient A., 35 years old, is presented with the diagnosis: Grade IIIA flame burn of the face, total body surface area involved = 5%. Expected outcomes of conventional treatment include a mean granulation formation period of approximately 21 days for Grade IIIA burn wounds and an average hospital stay of 30–35 days for patients with deep burns.
Burn treatment was performed using the Amplifier of Reparative Potentials for xenogeneic activation of autologous fibroblasts and keratinocytes. The working concentration was 1 million cells per milliliter (Figures 78 and 79).
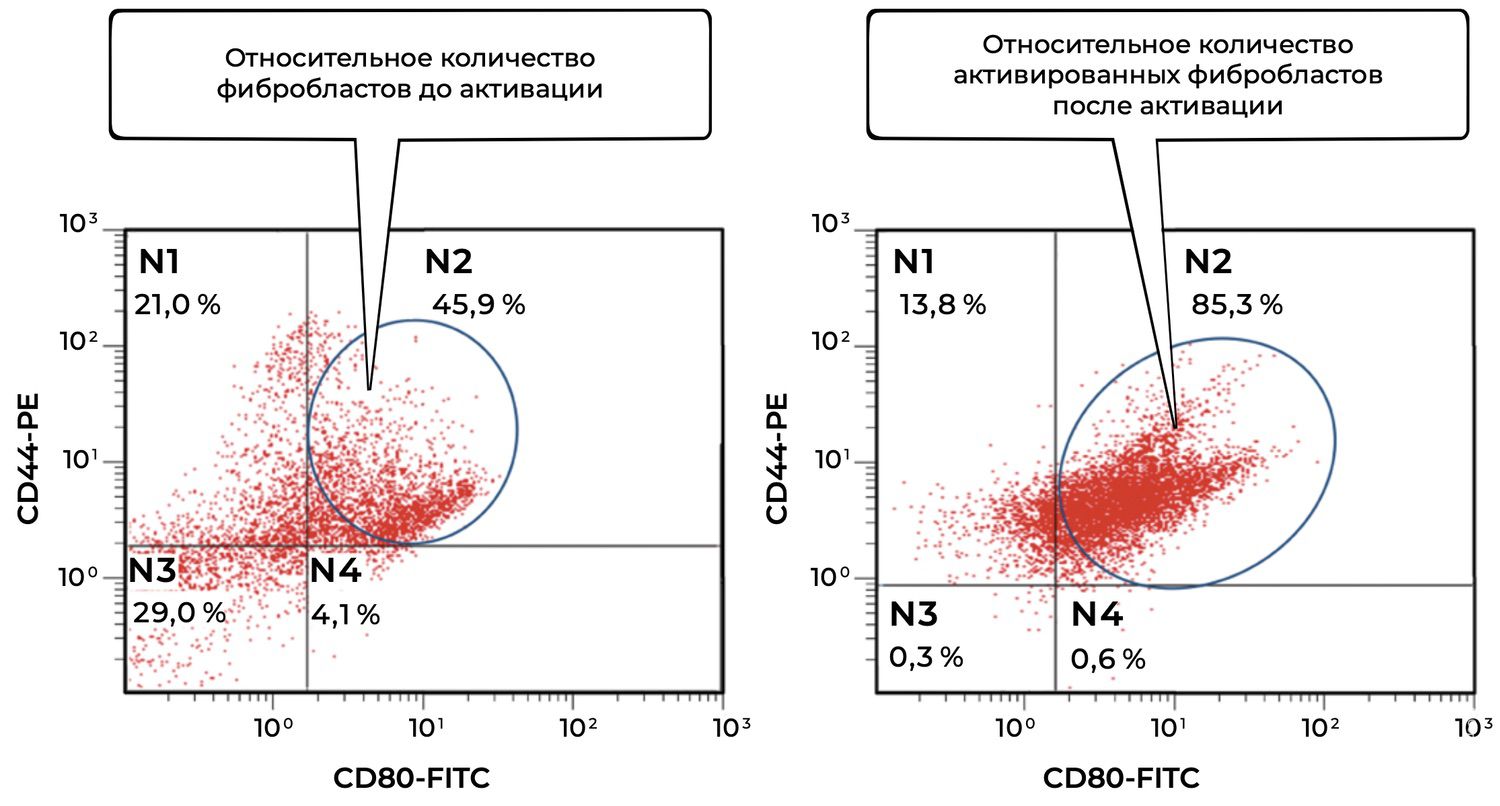
Figure 78. Comparative characteristics of the relative number of viable fibroblasts (CD45−CD14−CD44+ phenotype) before and after activation in the suspension of skin-biopsy cells obtained from Patient A., 35 years old, diagnosed with a Grade IIIA flame burn of the face.
The cytoimmunograms illustrate a comparative flow-cytometric analysis of CD80 and CD44 expression in dermal fibroblasts before and after activation.
The left panel demonstrates a cell population characterized by moderate CD44 expression and low CD80 expression:
Quadrant N2 (CD44⁺/CD80⁺): 45.9% — viable, partially activated fibroblasts.
Quadrant N1 (CD44⁺/CD80⁻): 21.0% — viable but non-activated fibroblasts.
Quadrant N3 (CD44⁻/CD80⁻): 29.0% — likely non-fibroblast cell populations.
Quadrant N4 (CD44⁻/CD80⁺): 4.1% — likely activated non-fibroblast cells.
The total proportion of viable fibroblasts was approximately 66.9%, a substantial fraction of which already exhibited signs of moderate activation (CD80⁺).
The right panel demonstrates a pronounced shift toward high CD80 expression:
Quadrant N2 (CD44⁺/CD80⁺): 85.3% — activated viable fibroblasts.
Quadrant N1 (CD44⁺/CD80⁻): 13.8% — non-activated fibroblasts.
Quadrant N3 (CD44⁻/CD80⁻): 0.3%.
Quadrant N4 (CD44⁻/CD80⁺): 0.6%.
Virtually the entire viable fibroblast population transitioned into an activated state, indicating the effectiveness of the activating agent.
Thus, activation resulted in a marked redistribution of fibroblasts toward a highly active phenotype, evidenced by more than a twofold increase in the proportion of CD44⁺CD80⁺ cells. These findings demonstrate:
a high intrinsic capacity of fibroblasts for in vitro activation;
the effectiveness of the activation procedure;
the potential value of cell-based burn therapy focused on fibroblast modulation.
Following activation, the proportion of fibroblasts co-expressing CD44 and CD80 increased from 45.9% to 85.3%, reflecting highly efficient stimulation of cellular activity. Simultaneous reduction of the N1, N3, and N4 populations confirms the targeted nature of activation and redistribution toward a functionally significant fibroblast phenotype. This finding may indicate substantial potential for stimulating dermal regeneration and enhancing cellular metabolism.
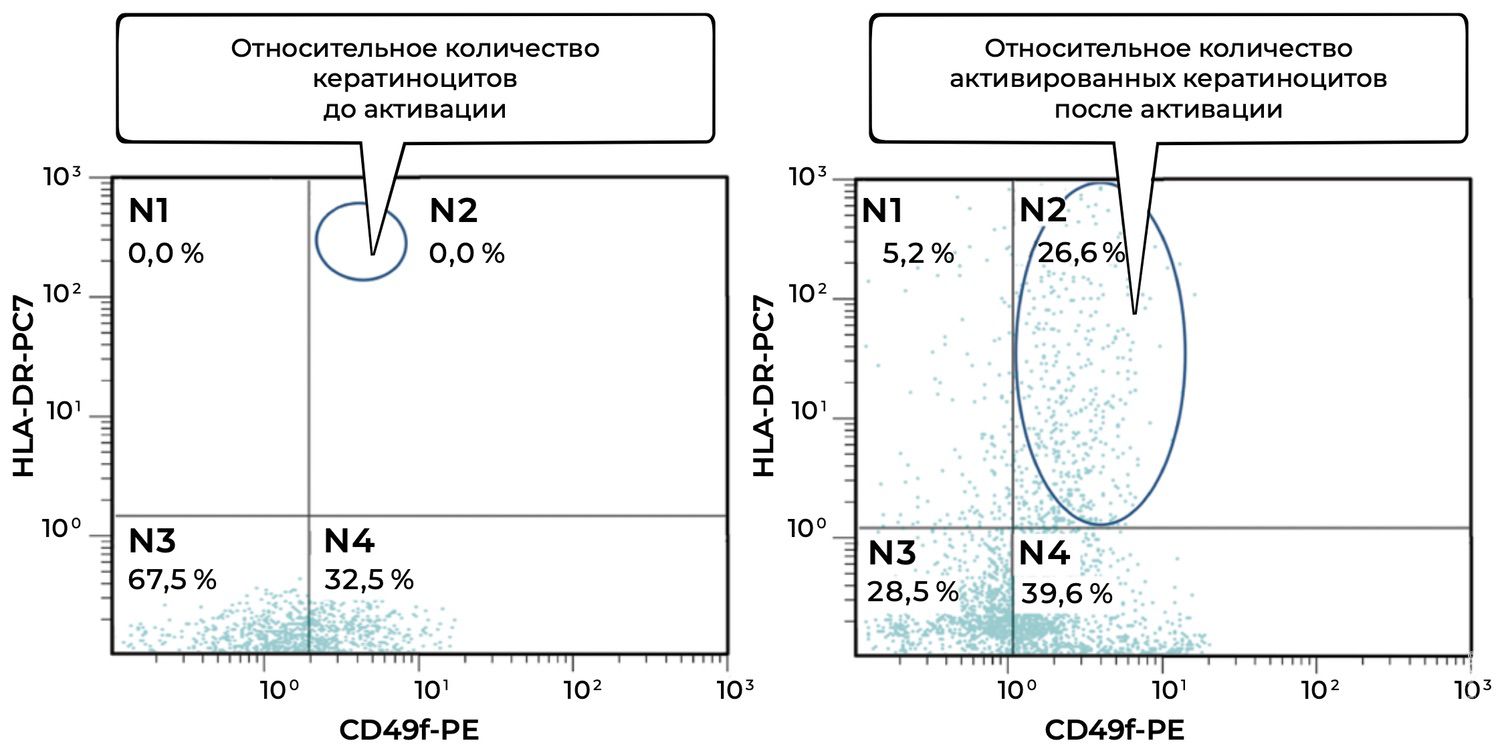
Figure 79. Comparative characteristics of the relative number of viable keratinocytes (CD49f⁺HLA-DR⁺ phenotype) before and after activation in the suspension of skin-biopsy cells obtained from Patient A., 35 years old, diagnosed with a Grade IIIA flame burn of the face.
Analysis of the cytoimmunograms provides a detailed characterization of changes in the viable keratinocyte population (CD49f⁺HLA-DR⁺ phenotype) before and after activation.
Quadrant N2 (CD49f⁺ / HLA-DR⁺): 0.0% — activated viable keratinocytes absent.
Quadrant N4 (CD49f⁺ / HLA-DR⁻): 32.5% — viable but non-activated keratinocytes.
Quadrants N1 and N3: 0.0% and 67.5%, respectively — cells lacking viability markers or not belonging to the keratinocyte lineage.
Keratinocytes were therefore almost entirely non-activated. The complete absence of CD49f⁺HLA-DR⁺ cells indicates profound suppression of epidermal regenerative responses.
Quadrant N2 (CD49f⁺ / HLA-DR⁺): 26.6% — substantial emergence of activated viable keratinocytes.
Quadrant N4 (CD49f⁺ / HLA-DR⁻): 39.6% — a proportion of cells remained viable but non-activated.
Quadrant N1: 5.2%.
Quadrant N3: 28.5%.
A marked increase in activated keratinocytes (CD49f⁺HLA-DR⁺) from 0% to 26.6% was observed, indicating restoration of epidermal proliferative potential.
Activation produced a clear shift toward recovery of epidermal function, demonstrated by the appearance of a substantial population of activated keratinocytes. These findings confirm the effectiveness of the activation procedure, indicate preservation of regenerative capacity within the patient's skin-cell population, and support the potential clinical utility of burn treatment strategies based on cellular modulation.
During treatment, 25 dressing changes were performed. Given the location, surface area, and depth of injury, sterile gauze dressings were applied after administration of the cell suspension containing activated keratinocyte and fibroblast cultures. Dressings were changed once daily (Figure 80). As a result of treatment, bright, well-vascularized granulation tissue developed within the burn wounds by day 15 after injury. General anesthesia was required only during the first and second procedures.
Figure 80. Clinical efficacy of fibroblast and keratinocyte cultures activated using the Amplifier of Reparative Potentials. Patient A., 35 years old.
Grade IIIA flame burn of the face, TBSA = 5%.
Formation of the granulation layer (day 10 after injury).
Fully developed granulation tissue and epithelialization of the lesion (day 20 after injury).
Final healing and restoration outcome (day 26 after injury).
The medical board noted significant advantages of using the Reparative Potential Activator in preparing cells for autotransplantation during burn treatment, particularly the reduction in epithelialization time, which shortened hospitalization by 10 days.
Ultimately, the clinical course observed in Patient A., 35 years old, following a Grade IIIA facial flame burn correlated closely with the cytoimmunogram findings.
Day of admission
Extensive destruction of facial and cervical skin.
Massive necrosis involving both epidermis and dermis.
Significant risk of scar formation and auricular deformity.
10 days after injury
Formation of granulation tissue indicating initiation of active regenerative processes.
Progressive wound cleansing and reduction of inflammation.
Active coverage of the wound surface with newly formed tissue.
20 days after injury
Granulation tissue undergoing epithelialization.
Areas of newly formed skin with restoration of texture and pigmentation.
Marked reduction of edema and hyperemia.
26 days after injury
Near-complete restoration of skin integrity.
Full preservation of the auricle without scar-related deformity.
Smooth, homogeneous skin texture indicating successful repair with minimal sequelae.
The cytoimmunogram findings, demonstrating activation of fibroblasts (up to 85.3%) and the emergence of activated keratinocytes (up to 26.6%), directly correlated with:
accelerated granulation-tissue formation and epithelialization;
substantial reduction in healing time and decreased risk of complications.
The medical board again emphasized the benefits of employing the Reparative Potential Activator in cell preparation for autotransplantation, particularly the shorter epithelialization period, which reduced hospitalization by ten days.
This clinical case serves as a compelling illustration of the effectiveness of activated autologous cell cultures (fibroblasts and keratinocytes) prepared using the Amplifier of Reparative Potentials. The combined laboratory and clinical findings demonstrate rapid restoration of skin integrity, regeneration with minimal scarring, and preservation of normal anatomical structures.
Patient S., 65 years old, is presented with the diagnosis: Grade IIB flame burn of the dorsum of the hand, total body surface area involved = 2%. Burn treatment was performed as monotherapy using the Amplifier of Reparative Potentials for xenogeneic activation of autologous keratinocytes. The working concentration was 1 million cells per milliliter (Figure 81).
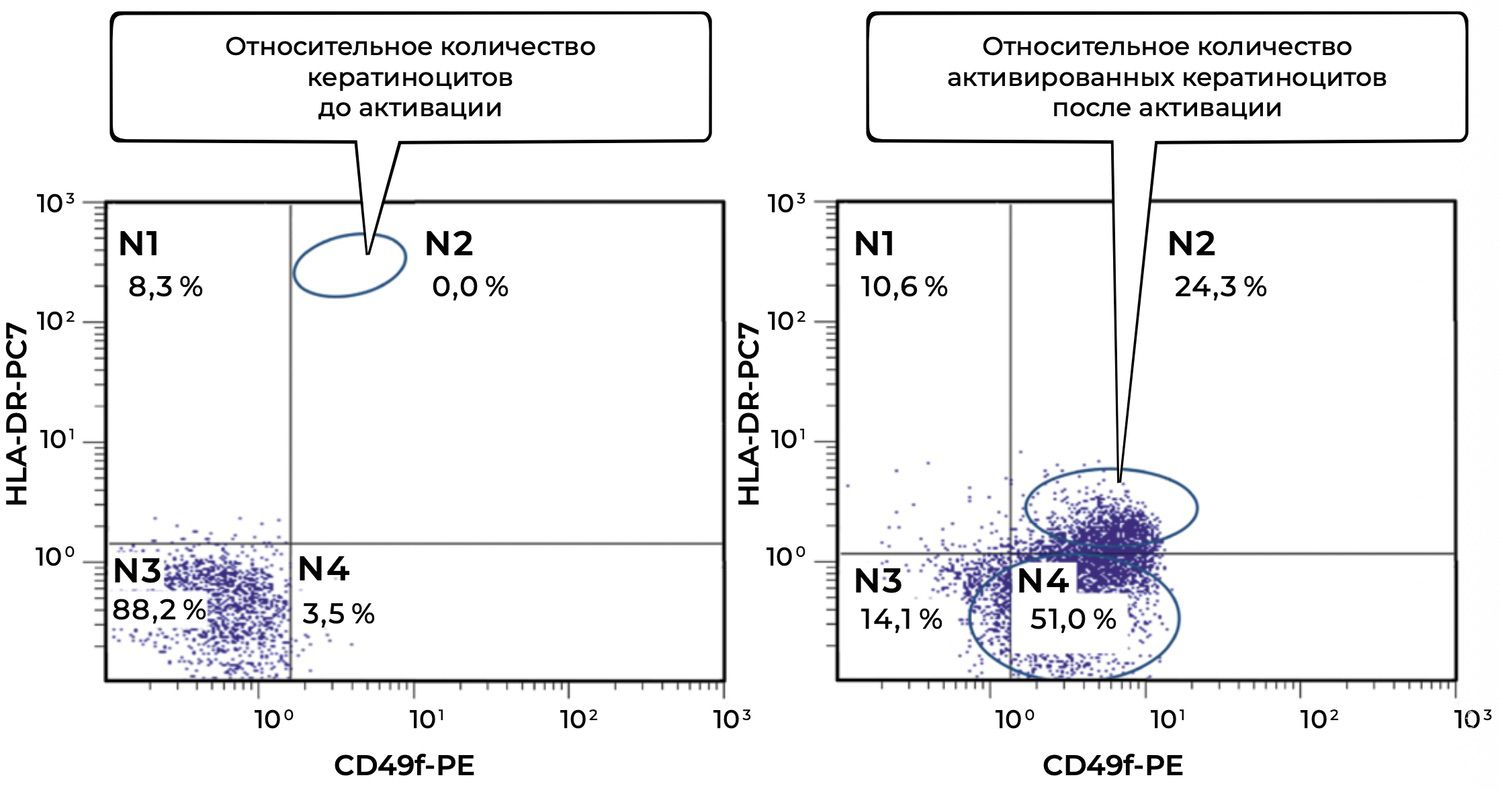
Figure 81. Comparative characteristics of the relative number of viable keratinocytes (CD49f⁺HLA-DR⁺ phenotype) before and after activation in the suspension of skin-biopsy cells obtained from Patient S., 65 years old, diagnosed with a Grade IIB flame burn of the dorsum of the hand.
The dynamics shown in the presented skin cytoimmunograms reflect changes in the relative proportion of viable keratinocytes (CD49f⁺HLA-DR⁺ phenotype) before and after activation in Patient S., 65 years old, diagnosed with a Grade IIB flame burn of the dorsum of the hand.
The relative proportion of activated keratinocytes (CD49f⁺HLA-DR⁺, sector N2) was 0.0%.
The majority of cells (88.2%) were localized in sector N3, corresponding to cells without CD49f and HLA-DR expression, possibly inactive or damaged cells.
Mature but non-activated keratinocytes (N4) accounted for 3.5%.
This pattern indicates a suppressed state of the epidermal compartment, with no signs of regenerative activity.
The relative proportion of activated keratinocytes (N2) increased significantly to 24.3%.
In parallel, the proportion of mature but non-activated keratinocytes (N4) increased to 51.0%.
There was a marked decrease in “non-specific” or potentially apoptotic cells (N3: 14.1% after activation versus 88.2% before activation).
Taken together, these data indicate restoration of cellular structure and initiation of epidermal regeneration, which is particularly important in superficial Grade IIB burns.
Thus, keratinocyte activation produced a distinct shift in the cytometric profile, manifested by:
dedifferentiation and reprogramming of part of the mature cell population;
emergence of a regenerative subpopulation expressing HLA-DR (N2);
reduction of “silent” or inactive cells.
Accordingly, in a burn of the dorsum of the hand in an elderly patient, induction of keratinocyte activation may be considered a critically important component of skin-barrier restoration. During the course of treatment, 14 dressing changes were performed once daily (Figure 82).
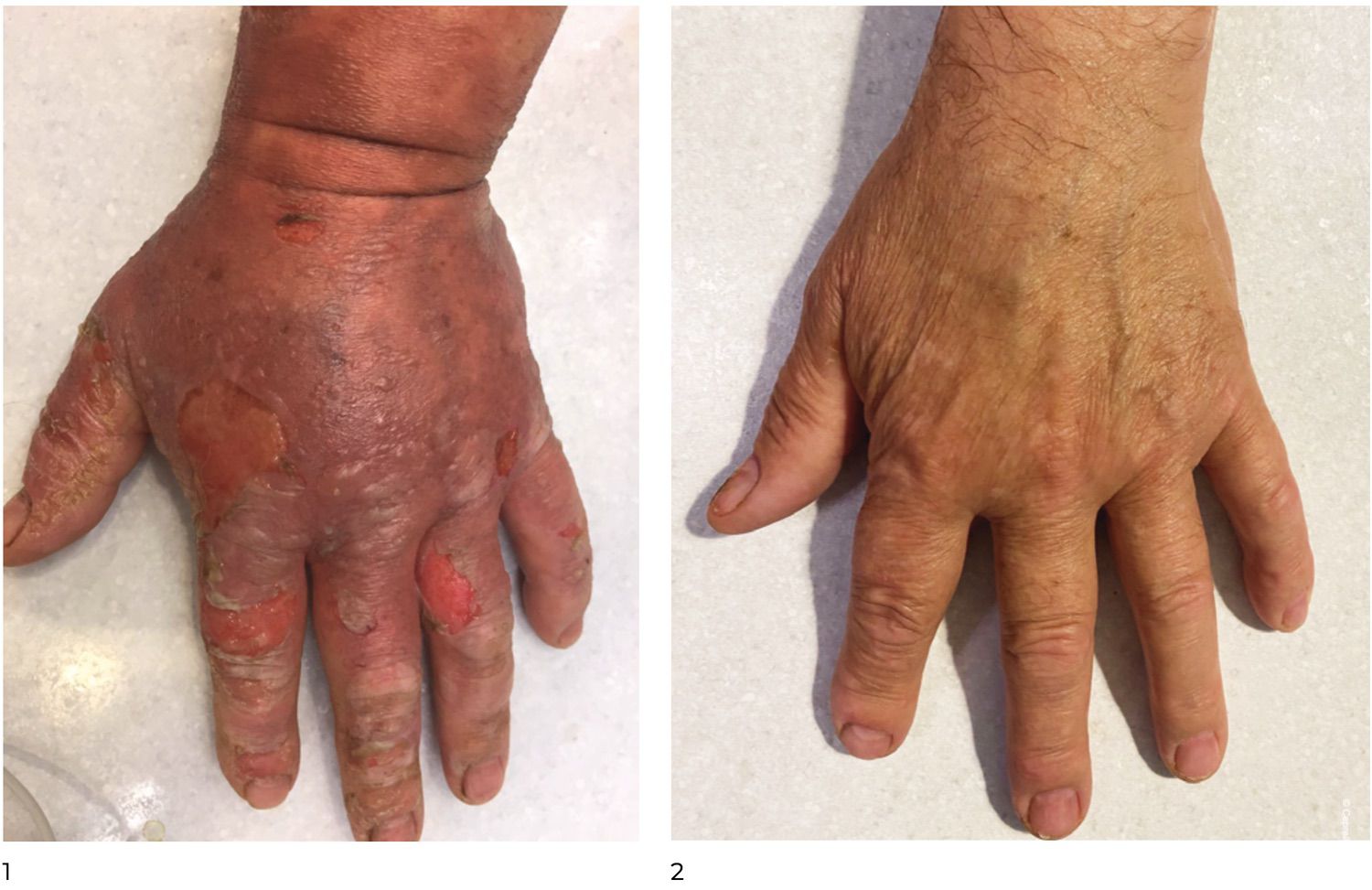
Figure 82. Clinical efficacy of a keratinocyte culture activated using the Amplifier of Reparative Potentials. Patient S., 65 years old, diagnosed with a Grade IIB flame burn of the dorsum of the hand.
Grade IIB flame burn of the dorsum of the hand.
Healing and restoration outcome on day 14 after injury.
This pair of photographs clearly demonstrates how laboratory cytoimmunological data correlate with clinical restoration of the skin in Patient S., 65 years old, with a Grade IIB thermal flame burn of the dorsum of the hand.
On the left, before treatment, there is pronounced inflammation, blistering, weeping erosions, and edema characteristic of a Grade IIB burn.
On the right, on day 14, complete epithelialization of the skin is observed, with resolution of edema, normalization of color and texture, and absence of coarse scarring.
| Parameter | Before treatment | After keratinocyte activation | Clinical result |
|---|---|---|---|
| CD49f⁺HLA-DR⁺ activated keratinocytes | 0.0% | 24.3% | Extensive erosions and edema → smooth, re-epithelialized skin surface |
| Overall keratinocyte viability | Low: 3.5% mature cells (CD49f⁺HLA-DR⁻) | Increased to 51.0% | Clear restoration of skin structure |
| Cytometric changes | Deficit of regenerative activity | Pronounced activation of epidermal cells | Successful restoration within 14 days |
The treatment outcome demonstrates the effectiveness of using activated keratinocyte cultures enhanced by the Amplifier of Reparative Potentials. Specifically, this approach:
significantly accelerates regeneration, even in elderly patients;
increases the functional activity of epidermal cells;
shortens healing time in thermal skin injuries.
This case demonstrates a direct relationship between immunocytological intervention and clinical effect, which is especially valuable for evaluating the efficacy of innovative burn therapy. It objectively confirms the effectiveness of applying the Activator of Reparative Potentials in preparing keratinocyte cultures for autotransplantation in burn treatment, namely shorter wound epithelialization time and a pronounced aesthetic result.
57 Гмурман, В. Е. Теория вероятностей и математическая статистика / В. Е. Гмурман. – Изд. 7-е, стер. – М.: Высшая школа, 2000. – 479 с.
58 Федеральные клинические рекомендации. Дерматовенерология 2015: Болезни кожи. Инфекции, передаваемые половым путем. – М. : Деловой экспресс, 2016. – 768 с.
59 Федеральные клинические рекомендации. Дерматовенерология 2015: Болезни кожи. Инфекции, передаваемые половым путем. – М. : Деловой экспресс, 2016. – 768 с.
60 Завгородняя, М. И. Клеточные и молекулярные основы заживления ран / М. И. Завгородняя, Л. В. Макеева, О. С. Славчева и др. // Morphologia. – 2016. – Т. 10. – № 3. – С. 19–23.
61 Li, J. Pathophysiology of acute wound healing / J. Li, J. Chen, R. Kirsner // Clinical Dermatology. – 2007. – Т. 25. – Р. 9–18.
62 Werner, S. Regulation of wound healing by growth factors and cytokines / S. Werner, R. Grose // Physiol. Rev. – 2003. – Т. 83. – Р. 835–870.
63 Алексеева, Н. Т. Роль клеток фибробластического дифферона в процессе заживления ран / Н. Т. Алексеева, А. А. Глухов, А. П. Остроушко // Вестник экспериментальной и клинической хирургии. – 2012. – Т. 5. – № 3. – С. 601–608.
64 Rittié, L. Cellular mechanisms of skin repair in humans and other mammals / L. Rittié // Journal of Cell Communication and Signaling. – 2016. – Т. 10 (2). – Р. 103–120.
65 Donati, G. Stem cell heterogeneity and plasticity in epithelia / G. Donati, F. M. Watt // Cell Stem Cell. – 2015. – Т. 16. – Р. 465–476.
66 Pal'tsev, M. A. Biologiya stvolovykh kletok i kletochnye tekhnologii / Pod red. M. A. Pal'tseva. – M.: Meditsina, 2009. –728 s.
67 Barinov, E. F. Histophysiology of hair follicles: current concept / E. F. Barinov, O. N. Sulaeva // Uspekhi fiziologicheskikh nauk. – 2003. – № 35 (4). – P. 65–77.
68 Rehman, J. Peripheral blood «endothelial progenitor cells» are derived from monocyte/macrophages and secrete angiogenic growth factors / J. Rehman, J. Li, C. M. Orschell et al. // Circulation. – 2003. – Т. 107. – Р. 1164–1169.
69 Chu, K. Distribution and in situ proliferation patterns of intravenously injected immortalized human neural stem-like cells in rats with focal cerebral ischemia / K. Chu, M. Kim, S. H. Chae et al. // Neuroscience Research. – 2004. – Т. 50 (4). – Р. 459–465.
70 Shintani, S. Augmentation of postnatal neovascularization with autologous bone marrow transplantation / S. Shintani, T. Murohara, H. Ikeda et al. // Circulation. – 2001. – Т. 103. – Р. 897–903.
71 Баринов, Э. Ф. Функциональная морфология кожи: от основ гистологии к проблемам дерматологии / Э. Ф. Баринов, Р. Ф. Айзятулов, М. Э. Баринова и др. // Клиническая дерматология и венерология. – 2012. – Т. 10. – № 1. – С. 90–93.
72 Kamihata, H. Implantation of bone marrow mononuclear cells into ischemic myocardium enhances collateral perfusion and regional function via side supply of angioblasts, angiogenic ligands, and cytokines / H. Kamihata, H. Matsubara, T. Nishiue et al. // Circulation. – 2001. – Т. 104. – Р. 1046–1052.
73 Donati, G. Stem cell heterogeneity and plasticity in epithelia / G. Donati, F. M. Watt // Cell Stem Cell. – 2015. – Т. 16. – Р. 465–476.
74 Терских, В. В. Стволовые клетки и структура эпидермиса / В. В. Терских, А. В. Васильев, Е. А. Воротеляк // Вестник дерматологии и венерологии. – 2005. – № 3. – С. 11–15.
75 Биология стволовых клеток и клеточные технологии: В 2 т. / Под ред. М. А. Пальцева. – М.: Медицина, 2009. – 272 с.
76 Домнина, А. П. Мезенхимальные стволовые клетки эндометрия человека при длительном культивировании не подвергаются спонтанной трансформации / А. П. Домнина, И. И. Фридлянская, В. И. Земелько и др. // Цитология. – 2013. – № 55 (1). – С. 69–74.
77 Приказ Министерства здравоохранения Российской Федерации от 19.01.2017 № 14н «Об утверждении формы спецификации на биомедицинский клеточный продукт».
78 Козлов, И. Г. Биоподобные препараты: в начале проблемы / И. Г. Козлов // Consilium Medicum. – 2010. – Т. 12. – № 12. – C. 94–98.
79 Сабурина, И. Н. Сопоставление поведения стромальных клеток пупочного канатика и мультипотентных стромальных клеток взрослого костного мозга в 2-d и 3-d-культуре: моделирование стромальной регенерации / И. Н. Сабурина, А. А. Горкун, Н. В. Кошелева, М. Л. Семенова, А. А. Пулин, В. С. Репин // Вестник новых медицинских технологий. – 2009. – Vol. 16. – № 4. – P. 9–11.
80 Antoni, D. Three-dimensional cell culture: a breakthrough in vivo / D. Antoni, H. Burckel, E. Josset, G. Noel // International Journal of Molecular Sciences. – 2015. – Vol. 16. – P. 5517–5527.
81 Moscona, A. A. The dissociation and aggregation of cells from organ rudiments of the early chick embryos / A. A. Moscona, H. Moscona // Journal of Anatomy. – 1952. – Vol. 86. – P. 287–303.
82 Moscona, A. A. Tissues from dissociated cells / A. A. Moscona // Scientific American. – 1959. – Vol. 200. – P. 132–134.
83 Steinberg, M. S. Reconsttution of tissues by dissociated cells: some morphogenetic tissue movements and the sorting out of embryonic cells may have a common explanation / M. S. Steinberg // Science. – 1963. – Vol. 141. – P. 401–408.
84 Steinberg, M. S. Differential adhesion and morphogenesis: a modern view / M. S. Steinberg // Curr. Opin. Genet. Dev. – 2007. – Vol. 17. – P. 281–285.
85 Сабурина, И. Н. Зd-культивирование: от отдельных клеток к регенерационной ткани (к вопросу о феномене эпителио-мезенхимальной пластичности) / И. Н. Сабурина, B.C. Репин // Гены и клетки. – 2010. – Vol. 5. – No. 2. – P. 75–86.
86 Манагаров, Н. Г. Костная пластика в челюстно-лицевой хирургии / Н. Г. Манагаров // Здравоохранение Югры: опыт и инновации. – 2018. – № 3 (16). – С. 27.
87 Довгилева, О. М. Основные особенности репаративной регенерации кожи в условиях применения хитозана / О. М. Довгилева, Г. В. Хомулло, М. Б. Петрова // Верхневолжский медицинский журнал. – 2011. – Т. 9. – Вып. 3. – № 11. – С. 30–37.
88 Li, H. W. Emerging concepts in haematopoietic cell transplantation / H. W. Li, M. Sykes // Nat. Rev. Immunol. – 2012. – Vol. 12 (6). – P. 403–416.
89 Бабаева, А. Г. Регенерация: факты и перспективы / А. Г. Бабаева. – М. : Издательство РАМН, 2009. – 336 с.
90 Ноздрин, В. И. Гистофизиология кожи / В. И. Ноздрин, С. А. Барашкова, В. В. Семченко. – Омск, 2008. – 280 с.
91 Данилов, Р. К. Раневой процесс: гистогенетические основы / Р. К. Данилов. – СПб. : Военно-медицинская академия имени С. М. Кирова, 2008. – 380 с.
92 Fuchs, Y. ARTS Regulates Stem Cell Apoptosis and Skin Regeneration / Y. Fuchs, S. Brown, T. Gorenc et al. // Science. – 2013. – Vol. 19. – No. 341 (6143). – Р. 286–289.
93 Engel, P. CD Nomenclature 2015: Human Leukocyte Differentiation Antigen Workshops as a Driving Force in Immunology / P. Engel et al. // J. Immunol. – 2015. – Т. 195 (10). – Р. 4555–4563.
94 Nery, A. A. Human Mesenchymal Stem Cells: From Immunophenotyping by Flow Cytometry to Clinical Applications / A. A. Nery, I. C. Nascimento, T. Glaser et al. // Cytometry. – 2013. – No. 83A. – Р. 48–61.
95 Donati, G. Stem cell heterogeneity and plasticity in epithelia / G. Donati, F. M. Watt // Cell Stem Cell. – 2015. – Т. 16. – Р. 465–476.
96 Козлов, И. Г. Биоподобные препараты: в начале проблемы / И. Г. Козлов // Consilium Medicum. – 2010. – Т. 12. – № 12. – C. 94–98.
97 Gibson, T. Zoografting – A curious chapter in the history of plastic surgery / T. Gibson // Br. J. of Plast. Surg. – 1955. – Vol. 8. – P. 234–242.
98 Prochorov, A. V. Long–term normalization of diabetes mellitus after xenotransplantation of fetal pancreatic islet cells into the blood stream without immunosuppressive therapy / A. V. Prochorov et al. // Transplantation Proceedings. – 2004. – Vol. 36. – P. 2855–2856.
99 Хрыщанович, В. Я. Ксеногенная клеточная терапия: современное состояние проблемы и перспективы свиной клеточной трансплантации / В. Я. Хрыщанович, С. И. Третьяк, А. А. Глинник // Журнал ГрГМУ. – 2012. – № 1 (37). – С. 14–23.
100 Fisher, M. B. Tissue engineering and regenerative medicine: recent innovations and the transition to translation / M. B. Fisher, R. L. Mauck // Tissue. Eng. Part B. Rev. – 2013. – V. 19. – No. 1. – P. 1–13.
101 Uludag, H. At the Intersection of Biomaterials and Gene Therapy: Progress in Non-viral Delivery of Nucleic Acids / H. Uludag, A. Ubeda, A. Ansari // Front Bioeng. Biotechnol. – 2019. – Т. 7. – Р. 131.
102 Fuchs, Y. ARTS Regulates Stem Cell Apoptosis and Skin Regeneration / Y. Fuchs, S. Brown, T. Gorenc et al. // Science. – 2013. – Vol. 19. – No. 341 (6143). – Р. 286–289.
103 Chang, S. E. Sphingosine may have cytotoxic effects via apoptosis on the growth of keloid fibroblasts / S. E. Chang, K. J. Kim, K. H. Ro // Journal of Dermatology. – 2004. – Vol. 31. – P. 1–5.
104 Herrmann, M. A rapid and simple method for the isolation of apoptotic DNA fragments / M. Herrmann, H.-M. Lorenz, R. Voll, M. Grunke, W. Woith, J. R. Kalde // Nucleic Acids Research. – 1994. – Vol. 22. –P. 5506–5507.
105 Sato, J. D. Media for culture of mammalian cells / J. D. Sato, M. Kan // Current Protocols in Cell Biology. –2001. – Vol. 1. – P. 12–15.
106 Zheng, X. Proteomic analysis for the assessment of different lots of fetal bovine serum as a raw material for cell culture. Part IV. Application of proteomics to the manufacture of biological drugs / X. Zheng, H. Baker, W. S. Hancock, F. Fawaz, M. McCaman, E. Pungor // Biotechnology progress. – 2006. – Vol. 22. – No 5. – P. 1294–1300.
107 Надеев, А. Д. Индукция апоптоза и некроза клеток эндотелия пупочной вены человека пероксидом водорода / А. Д. Надеев, И. В. Кудрявцев, М. К. Серебрякова и др. // Цитология. – 2015. – Т. 57. – № 12. – С. 909–916.
108 Tallarida, R. J. Drug synergism and dose-effect data analysis / R. J. Tallarida. – CRC Press, 2000. – 264 p.
109 Khuri, A. I. Response surface methodology / A. I. Khuri, S. Mukhopadhyay // Wiley Interdisciplinary Reviews: Computational Statistics. – 2010. – Т. 2. – No. 2. – Р. 128–149.
110 Костоломова, Е. Г. Некоторые иммунофизиологические механизмы регенерации ран в условиях применения ранозаживляющего средства Cellgel / Е. Г. Костоломова, Ю. Г. Суховей, С. В. Гольцов и др. // Российский иммунологический журнал. – 2016. – № 10 (19). – С. 289–291.
111 Гистологическая техника : учеб. пособие / под ред. В. В. Семченко, С. А. Барашкова, В. Н. Ноздрина и др. – Омск; Орел : Омская областная типография, 2006. – 290 с.
112 Попова, Л. Н. Как изменяются границы вновь образующегося эпидермиса при заживлении ран : автореф. дис. … канд. мед. наук / Л. Н. Попова. – М., 1942. – 16 с.
113 Шарафутдинова, И. Р. Инновационные технологии в мониторинге скорости заживления ран / И. Р. Шарафутдинова, З. З. Мустафина, А. Я. Габитова и др. // Международный студенческий научный вестник. – 2018. – № 4. – Ч. 1. – С. 177–179.
114 Патент № 2481115 Российская Федерация, Средство для заживления ран «Cellgel», способ его получения и способ лечения ран различной этиологии полученным средством / С. В. Гольцов, Ю. Г. Суховей, Е. Г. Костоломова и др.; патентообладатель Гольцов С. В. – № 2011141622/15 заявл. 13.10.2011; опубл. 10.05.2013, Бюл. № 13.
115 Гольцов, С. В. Cellgel – новое слово в ранозаживлении / С. В. Гольцов, Е. Н. Гольцова, А. Д. Гетьман и др. // Дерматология в России. – 2017. – № 1. – С. 28–33.
116 Костоломова, Е. Г. Некоторые иммунофизиологические механизмы регенерации ран в условиях применения ранозаживляющего средства Cellgel / Е. Г. Костоломова, Ю. Г. Суховей, С. В. Гольцов и др. // Российский иммунологический журнал. – 2016. – № 10 (19). – С. 289–291.
117 Костоломова, Е. Г. Роль процессов пролиферации и апоптоза в образовании рубцовой ткани / Е. Г. Костоломова, С. А. Стрелин, Ю. Г. Суховей и др. // Медицинская иммунология. – 2017. – № 19 (5). – С. 327.
118 Костоломова, Е. Г. Взаимодействие иммуноцитов кожи в процессе репаративной регенерации в ране / Е. Г. Костоломова, Ю. Г. Суховей, И. Г. Унгер и др. // Российский иммунологический журнал. – 2017. – № 2 (20). – С. 148–150.
119 Суховей, Ю. Г. Регенераторно-репаративные и антибактериальные свойства препарата Cellgel в эксперименте / Ю. Г. Суховей, Е. Г. Костоломова, С. Б. Цирятьева и др. // Российский иммунологический журнал. – 2015. – № 9 (2). – С. 44.
120 Патент № 159463 Российская Федерация, Устройство для активации репаративных потенций клеток кожи / С. В. Гольцов, Ю. Г. Суховей, Е. Г. Костоломова и др.; патентообладатель Гольцов С. В. – № 2015131577/10; заявл. 29.07.2015; опубл. 10.02.2016, Бюл. № 4.
121 Петрова, Н. П. Система независимой оценки (подтверждения) соответствия парфюмерно-косметической продукции в Российской Федерации / Н. П. Петрова, А. В. Резайкина, С. В. Ротанов // Вестник дерматологии и венерологии. – 2015. – № 6. – С. 90–96.
122 Кубанова, А. А. Перспективы практического использования наночастиц в дерматологии / А. А. Кубанова, С. Р. Утц, А. А. Кубанов и др. // Вестник дерматологии и венерологии. – 2016. – № 2. – С. 15–20.
123 Тараховский, Ю. С. Интеллектуальные липидные наноконтейнеры в адресной доставке лекарственных веществ / Ю. С. Тараховский. – М. : ЛКИ, 2011. – 280 с.
124 Papakostas, D. Nanoparticles in dermatology / D. Papakostas, F. Rancan, W. Sterry et al. // Archives of Dermatological Research. – 2011. – Т. 303. – Р. 533–550.
125 Воротеляк, Е. А. Организация популяции кератиноцитов в культуре in vitro / Е. А. Воротеляк, Э. С. Чермных, А. В. Васильев и др. // Известия РАН. Сер. биол. – 2005. – № 6. – С. 645–649.
126 Костоломова, Е. Г. Некоторые иммунофизиологические механизмы регенерации ран в условиях применения ранозаживляющего средства Cellgel / Е. Г. Костоломова, Ю. Г. Суховей, С. В. Гольцов и др. // Российский иммунологический журнал. – 2016. – № 10 (19). – С. 289–291.
127 Воротеляк, Е.А. Регенерация кожи: очевидные модели и неочевидные результаты / Е. А. Воротеляк, Е. И. Моргун, Э. С. Чермных, О. С. Роговая, Е. П. Калабушева // Гены и клетки. – 2022. – Т. 17. – № 3. – С. 47–47.
к предыдущей странице | читать далее | сразу к следующей главе



